Liaoningvenator - Liaoningvenator
| Liaoningvenator | |
|---|---|
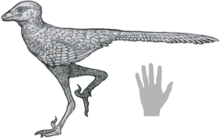 | |
| Реконструкция жизни рукой за масштаб | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Troodontidae |
| Род: | †Liaoningvenator Шен и другие., 2017 |
| Типовой вид | |
| †Liaoningvenator curriei Шен и другие., 2017 | |
Liaoningvenator (смысл "Ляонин охотник ") род из троодонтид теропод динозавр от Раннемеловой период из Китай. Он содержит один вид, L. curriei, названный в честь палеонтолог Филип Дж. Карри в 2017 году Шен Кай-Чжи и его коллеги из сочлененного, почти полного скелета, одного из наиболее полных известных экземпляров троодонтид. Шен и его коллеги обнаружили характерные черты, которые Liaoningvenator внутри Troodontidae. Эти черты включали его многочисленные маленькие и плотно прилегающие зубы, а также позвонки ближе к концу хвоста, имеющие мелкие бороздки вместо нервные отростки на их верхних поверхностях.
В пределах Troodontidae ближайший родственник Liaoningvenator был Эозиноптерикс, и это также было тесно связано с Анхиорнис и Сяотия; в то время как они традиционно размещались за пределами Troodontidae, филогенетический анализ Шен и его коллеги предоставили доказательства, подтверждающие альтернативную идентификацию этих паравианцы как троодонтиды. Однако по сравнению со своими близкими родственниками Liaoningvenator имели относительно длинные ноги, в частности бедра. Как пятый троодонтид, известный из географически и временно сопоставимых слои, Liaoningvenator увеличивает разнообразие китайских троодонтид.
Описание
При общей длине тела (предполагаемая длина черепа, шеи, туловища и хвоста вместе взятых) примерно 69 см (27 дюймов), Liaoningvenator это маленький троодонтид.[1]
Череп и челюсти
Liaoningvenator обладает длинным, узким и треугольным черепом длиной 9,8 см (3,9 дюйма). В передней части морды, как Синовенатор и Xixiasaurus, то предчелюстная кость исключает верхняя челюсть от края ноздри. На поверхности верхней челюсти есть три отверстия: предчелюстная, верхнечелюстная и верхнечелюстная. анторбитальные отверстия. Внизу верхняя челюсть образует вторичное небо как в Байронозавр, Гобивенатор, и Xixiasaurus. Уникальный среди троодонтид заглазничная кость тонкий и излучается на три отростка. Нравиться Занабазар, Существует пневматический дивертикул в скуловая кость там, где в кости находился воздушный мешок; есть также пневматическое отверстие на задней стороне квадратная кость, как и у других троодонтид.[2] В отличие от Sauronithoides, Занабазар, и Стенонихозавр, гребень, разделяющий теменные кости не участвует в границе надвисочного (верхнего) височного отверстия на задней части черепа.[1]
Характеристика троодонтид,[2] Liaoningvenator имеет выемчатую канавку на внешнем крае мелкой треугольной нижней челюсти. Нижний край челюсти слегка выпуклый; в Sinornithoides, прям. В зубной и угловатые кости могли образовать гибкий сустав внутри челюсти, то есть внутринижнечелюстной сустав.[3] В отличие от Сяотия дентальная и верхняя челюсти оканчиваются в одном и том же месте челюсти. Также как и другие троодонтиды (с Синусоназус являясь исключением), Liaoningvenator имеет ряд маленьких, близко расположенных зубов, по крайней мере 15 в верхней челюсти и 23 в нижней челюсти. Зубы по направлению к задней части нижней челюсти зазубрены, в отличие от ряда базальный троодонтиды. Зубной ряд верхней челюсти заканчивается ниже переднего края анторбитального отверстия, в то время как он заканчивается дальше вперед - ниже задней части верхнечелюстного отверстия. Джинфенгоптерикс.[1]
Позвонки
Есть десять шейные позвонки, двенадцать спинные (туловищные) позвонки, и не менее шестнадцати хвостовых (хвостовых) позвонков в Liaoningvenator. Из шейных позвонков удлиненные с третьего по восьмой, причем пятая - самая длинная; среди суставные отростки Пятая шейка матки, известная как презигапофизы, также имеет наибольшую длину. В третьем и четвертом шейных отделах последний, как и другие полученный troodontids презигапофизы длиннее, чем другой набор отростков, известный как постзигапофизы. В дорсальной части пневматическая точечная обработка упрощена по сравнению с Анхиорнис. В хвосте точка перехода - точка, где стороны хвостовых костей становятся более сжатыми, так что они становятся субтреугольными, а не прямоугольными, - происходит на седьмом хвосте, дальше вперед, чем Sinornithoides и Мэй (где это происходит на девятом).[4][5] Самая длинная из хвостовых костей - четырнадцатая, что почти в два раза длиннее шестой. На нижней стороне хвостовых костей шевроны слегка изогнуты и направлены назад, как это видно на Дейноних.[1]
Конечности
В акромион процесс на лопатка из Liaoningvenator слабо развит, как у базальных троодонтид. Однако в отличие от базальных троодонтид гленоид - гнездо для руки - направлено вертикально вниз, а не в сторону. На плечевая кость дельто-грудной гребень простирается на 40% длины кости и заканчивается на полпути вниз по кости. Плечевая кость короче по сравнению с бедренная кость, в Liaoningvenator (59% длины), чем в Эозиноптерикс (80% длины).[6] Ниже трехзначная стрелка уникальна тем, что первая фаланга первой цифры длиннее второй пястный, в 1,49 раза длиннее последнего.[1]
Как в Мэй, верхняя часть подвздошная кость имеет изогнутую синусоидальную форму в Liaoningvenator. Нет гребня над вертлужная впадина (тазобедренный сустав) из Liaoningvenator, В отличие от Анхиорнис. Задняя часть подвздошной кости (постацетабулярный отросток) также имеет более короткий нижний край, чем оба Анхиорнис и Эозиноптерикс. Уникально то, что на верхнем конце кончика седалищная кость, а кость также имеет тонкую запирательный процесс. В лобок указывает вперед в Liaoningvenator, но в обратном направлении Мэй.[5] Кроме того, задняя конечность Liaoningvenator вдвое длиннее туловища, а в Мэй это в 2,8 раза больше длины туловища. В отличие от Синовенатор, Liaoningvenator имеет четвертый вертел на бедренной кости. В большеберцовая кость тонкий и в 1,4 раза длиннее бедра, как и Sinornithoides. Стопа с четырьмя пальцами сильно уплотнена, с предплюсна который сужается к низу. Третий плюсневой смещен от второго и четвертого, образуя впадину между двумя последними, более глубокую, чем у других троодонтид. Пропорционально первая фаланга второго пальца короче второй фаланги у Liaoningvenator (135% длины), чем в Синовенатор (150% длины).[1][7]
Открытие и наименование
Liaoningvenator известен по единственному экземпляру, почти полному и хорошо сохранившемуся скелету с большинством костей, сохранившихся в их первоначальных сочлененных положениях. Он был найден в пластах Луцзятунь Формация Исянь в Шанъюане, Бейпяо, Ляонин, Китай; в настоящее время он хранится в Даляньском музее естественной истории (DNHM) в Далянь, Ляонин под инвентарным номером DNHM D3012. Некоторые из костей морды экземпляра неполные, а некоторые фаланги пальцев были добавлены незаконными торговцами окаменелостями; помимо этого, это одна из наиболее полных окаменелостей троодонтид, когда-либо обнаруженных. Его голова наклонена вперед, а конечности сведены внутрь; это отличается как от классического поза смерти (где голова наклонена назад), а также поза Мэй и Sinornithoides.[1]
В 2017 году DNHM D3012 был назван типовой образец нового рода и вида Liaoningvenator curriei Шэнь Цайчжи, Чжао Бо, Гао Чуньлин, Люй Цзюньчан и Мартин Кундрат. Название рода Liaoningvenator сочетает Ляонин с суффиксом -венатор, что означает "охотник" в латинский; конкретное имя Curriei отмечает вклад Канадский палеонтолог Филип Джон Карри к изучению мелких теропод.[1]
Классификация
Шен и его коллеги определили Liaoningvenator как член Troodontidae из-за его многочисленных, близко расположенных зубов, суженных ниже Корона; пневматическое отверстие в задней части его квадрата; овальная форма его большое затылочное отверстие; замена нервные отростки неглубокими срединными бороздками на позвонках ближе к концу хвоста; высокий восходящий процесс на его астрагал; и его асимметричный и субарктометатарзальный (то есть там, где третья плюсневая кость несколько ущемлена соседними плюсневыми костями) стопы.[2][8] Они также поместили его в «кладу высших троодонтид» из-за отсутствия луковичной капсулоподобной структуры на парасфеноид своего нёбо и наличие на черепе переднечелюстного отверстия.[1]
На основе филогенетический анализ модифицированный на основе предыдущего анализа Таканобу Цуйхиджи и его коллег в 2016 году, который, в свою очередь, был изменен на основе модификации Гао и его коллег в 2012 году анализа, проведенного Сюй Син и коллег в 2012 году,[9] Шен и его коллеги обнаружили, что Liaoningvenator образовали единую группу, или клады, с Эозиноптерикс, Анхиорнис, и Сяотия, предлагая тем самым свидетельство противоположного традиционному размещению этих таксоны как нетроодонтидные члены Паравес. Они объединены тем, что зубы уплощены и загнуты назад, а высота коронок в середине зубного ряда меньше, чем в два раза их ширины; передний край акромиона вывернут наружу; наличие ярко выраженной выемки между акромионом и коракоидный; наличие фланца на первой фаланге второго пальца пальца; и выпуклый лобок.[1]
Внутри этой клады, которую Шен и его коллеги не назвали, Liaoningvenator сформировал группу с Эозиноптерикс пока Анхиорнис сформировал группу с Сяотия. Первые два объединяет отсутствие зубчатых зубов на передней части челюсти; длина черепа превышает 90% длины бедренной кости; шейные ребра имеют тонкие стержни и длиннее соответствующих позвонков; и передний конец подвздошной кости «слегка прямой». Между тем, последние два имеют веерообразную веерообразную форму кончиков нервных шипов на спине; коракоид подтреугольный; коготь на первом пальце кисти сильно выгнут, выше вершины суставной поверхности; наличие «губки» на верхнем конце когтей на втором и третьем пальцах; передний край лобкового вала выпуклый; когти на третьем и четвертом пальцах стопы сильно изогнуты. Результаты филогенетического анализа воспроизведены ниже. филогенетическое дерево.[1]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Гистология
Тонкие секции из большеберцовая кость из голотип образец Liaoningvenator указать, что кортикальная кость имеет толщину 1,5 мм (0,059 дюйма). Кора делится на четыре зоны: линии задержанного роста (LAG), что указывает на то, что животному было не менее четырех лет, когда оно умерло (дальнейшие LAG могли быть уничтожены расширением мозговая полость ). Шен и его коллеги предположили, исходя из толщины самой внутренней зоны 1 по сравнению с зоной 2, что она частично разрушилась. Однако вся зона 3 еще тоньше, составляя лишь четверть ширины зоны 2, а зона 4 еще тоньше (хотя и неполная). Это указывает на то, что к концу третьего года рост существенно замедлился.[1]
Каждый LAG окружен двумя полосами плотной бессосудистой (то есть без отверстий для кровеносных сосудов) кости, которые Шен и его коллеги назвали «пре-кольцевыми» и «пост-кольцевыми» полосами. Первый LAG отличается от других тем, что он состоит из двух LAG, один из которых слабее другого, что указывает на незначительное замедление роста до возобновления роста костей. На внешнем крае кости нет внешней фундаментальной системы (EFS), что указывает на то, что голотип все еще рос на момент смерти. Однако уменьшение толщины зон, наличие бессосудистой кости во внешних слоях и свидетельства ремоделирование костей в совокупности предполагают, что он был близок к зрелости скелета.[1]
Палеоэкология
По словам Шена и его коллег, Liaoningvenator один из одиннадцати троодонтид, известных из Китая, и пятый раннемеловой китайский троодонтид после Синовенатор, Синусоназус, Мэй, и Джинфенгоптерикс.[1] В отдельной публикации 2017 года, в которой Шен также был ведущим автором, был описан дополнительный троодонтид, Далианзавр, который формирует Sinovenatorinae с бывшими тремя.[10] За исключением Джинфенгоптериксвсе эти троодонтиды обитали в слоях луцзятунь. В то время как Шен и его коллеги отнесли кровати Люцзятунь к Готеривский эпохи, более новые оценки даты, опубликованные Чанг Су-чин и его коллеги, предполагают более молодой возраст ~ 126 лет. Ма для пластов Луцзятунь, датируемых Барремский эпоха.[11]
Современные динозавры включали микрорапторин дромеозаврид Грацилираптор; то овирапторозавр Инцизивозавр; то орнитомимозавры Шэньчжоузавр[12] и Проклятье;[13] то процедурозаврид тираннозавроид Дилонг;[12] то титанозавр зауропод Euhelopus;[14] то орнитопод Jeholosaurus; и цератопсы такие как вездесущие Пситтакозавр[15] а также Ляоцератопс.[12] Присутствующие млекопитающие включены Акристатерий,[16] Гобиконодон, Juchilestes, Маотерий, Meemannodon, и Репеномам.[17][18][19] Другой четвероногие включены лягушки Ляобатрахус[20] и Мезофрин;[21] и ящерица Далингозавр.[22] Кровати Lujiatun состоят из речной и вулканический отложения, указывающие на ландшафт рек, несущих вулканы,[1] который мог убить сохранившихся животных лахар.[19] Среднегодовая температура воздуха в регионе достигла минимум 10 ° C (50 ° F).[23]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о Shen, C.-Z .; Чжао, Б .; Gao, C.-L .; Lu, J.-C .; Кундрат, М. «Новый динозавр троодонтид (Liaoningvenator curriei ген. et sp. nov.) из раннемеловой свиты Исянь в провинции Западный Ляонин ». Acta Geoscientica Sinica. 38 (3): 359–371. Дои:10.3975 / cagsb.2017.03.06.
- ^ а б c Makovicky, P.J .; Норелл, М.А. (2004). "Troodontidae". In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 184–195.
- ^ Tsuihiji, T .; Barsbold, R .; Watabe, M .; Цогтбаатар, К .; Чинзориг, Т .; Fujiyama, Y .; Сузуки, С. (2014). «Прекрасно сохранившийся теропод троодонтид с новой информацией о небной структуре из верхнего мела Монголии». Naturwissenschaften. 101 (2): 131–142. Bibcode:2014NW .... 101..131T. Дои:10.1007 / s00114-014-1143-9. PMID 24441791.
- ^ Currie, P.J .; Донг, З. (2001). «Новые сведения о троодонтидах мелового периода (Dinosauria, Theropoda) из Китайской Народной Республики». Канадский журнал наук о Земле. 38 (12): 1753–1766. Дои:10.1139 / e01-065.
- ^ а б Gao, C .; Morschhauser, E.M .; Varricchio, D.J .; Liu, J .; Чжао, Б. (2012). "Второй крепко спящий дракон: новые анатомические детали китайского троодонтида" Мэй Лонг с последствиями для филогении и тафономии ". PLOS ONE. 7 (9): e45203. Дои:10.1371 / journal.pone.0045203. ЧВК 3459897. PMID 23028847.
- ^ Godefroit, P .; Demuynck, H .; Дайк, G .; Hu, D .; Escuillié, F.O .; Клэйс, П. (2013). «Уменьшение оперения и летной способности нового юрского паравианского теропод из Китая». Nature Communications. 4: 1394. Bibcode:2013 НатКо ... 4,1394 г. Дои:10.1038 / ncomms2389. PMID 23340434.
- ^ Сюй, X .; Чжао, Цзи; Салливан, К .; Tan, Q.-W .; Сандер, М .; Ма, Q.-Y. (2012). «Пересмотрена таксономия троодонтид IVPP V 10597» (PDF). Позвоночные PalAsiatica. 50 (2): 140–150.
- ^ Тернер, A.H .; Makovicky, P.J .; Норелл, М.А. (2012). «Обзор систематики дромеозавров и паравианской филогении». Бюллетень Американского музея естественной истории. 371: 1–206. Дои:10.1206/748.1. HDL:2246/6352.
- ^ Tsuihiji, T .; Barsbold, R .; Watabe, M .; Цогтбаатар, К .; Suzuki, S .; Хаттори, С. (2016). «Новый материал теропода троодонтида (Dinosauria: Saurischia) из нижнего мела Монголии». Историческая биология. 28 (1–2): 128–138. Дои:10.1080/08912963.2015.1005086.
- ^ Shen, C .; Lü, J .; Liu, S .; Kundrát, M .; Brusatte, S.L .; Гао, Х. (2017). «Новый динозавр троодонтид из формации Исянь нижнего мела провинции Ляонин, Китай» (PDF). Acta Geologica Sinica. 91 (3): 763–780. Дои:10.1111/1755-6724.13307.
- ^ Chang, S.-C .; Gao, K.-Q .; Чжоу, З.-Ф .; Журдан, Ф. (2017). «Новые хроностратиграфические ограничения формации Исянь с последствиями для биоты Джехол». Палеогеография, палеоклиматология, палеоэкология. 487: 399–406. Bibcode:2017ППП ... 487..399С. Дои:10.1016 / j.palaeo.2017.09.026.
- ^ а б c Сюй, X .; Норелл, М.А. (2006). «Ископаемые останки динозавров, не относящихся к птицам, из нижнемеловой группы Джехол в западном Ляонине, Китай» (PDF). Геологический журнал. 41 (3–4): 419–437. Дои:10.1002 / gj.1044. Архивировано из оригинал (PDF) на 2016-03-03. Получено 2017-12-03.
- ^ Jin, L .; Jun, C .; Годфройт, П. (2012). «Новый базальный орнитомимозавр (Dinosauria: Theropoda) из раннемеловой формации Исянь, Северо-Восточный Китай» (PDF). В Godefroit, P. (ed.). Динозавры Берниссара и наземные экосистемы раннего мела. Жизнь прошлого. Блумингтон: Издательство Индианского университета. С. 467–487. ISBN 978-0-253-35721-2.
- ^ Barrett, P.M .; Ван, X.-L. (2007). "Базальные зубы титанозавров (Dinosauria, Sauropoda) из нижнемеловой формации Исянь провинции Ляонин, Китай". Палеомир. 16 (4): 265–271. Дои:10.1016 / j.palwor.2007.07.001.
- ^ Hedrick, B.P .; Додсон, П. (2013). «Пситтакозавриды Луцзятун: понимание индивидуальных и тафономических вариаций с использованием трехмерной геометрической морфометрии». PLOS ONE. 8 (8): e69265. Bibcode:2013PLoSO ... 869265H. Дои:10.1371 / journal.pone.0069265. ЧВК 3739782. PMID 23950887.
- ^ Hu, Y .; Meng, J .; Li, C .; Ван, Ю. (2010). «Новое базальное эвтериальное млекопитающее из раннемеловой биоты Джехол, Ляонин, Китай». Труды Королевского общества B. 277 (1679): 229–236. Дои:10.1098 / rspb.2009.0203. ЧВК 2842663. PMID 19419990.
- ^ Wang, Y.-Q .; Ху, Ю.-М .; Лизать. (2006). «Обзор последних достижений в изучении мезозойских млекопитающих в Китае» (PDF). Позвоночные PalAsiatica. 44 (2): 193–204.
- ^ Лопатин, А .; Аверьянов, А. (2015). "Гобиконодон (Mammalia) из раннего мела Монголии и ревизия Gobiconodontidae ». Журнал эволюции млекопитающих. 22 (1): 17–43. Дои:10.1007 / s10914-014-9267-4.
- ^ а б Jiang, B .; Fürsich, F.T .; Sha, J .; Ван, Б .; Ню, Ю. (2011). «Раннемеловой вулканизм и его влияние на сохранение окаменелостей в Западном Ляонине, северо-восток Китая». Палеогеография, палеоклиматология, палеоэкология. 302 (3): 255–269. Дои:10.1016 / j.palaeo.2011.01.016.
- ^ Dong, L .; Roček, Z .; Wang, Y .; Джонс, M.E.H. (2013). «Незубые животные из нижнемеловой группы Джехол в Западном Ляонине, Китай». PLOS ONE. 8 (7): e69723. Дои:10.1371 / journal.pone.0069723. ЧВК 3724893. PMID 23922783.
- ^ Wang, Y .; Jones, M.E.H .; Эванс, С. (2007). «Молодой ануран из формации Цзюфотан нижнего мела, Ляонин, Китай» (PDF). Меловые исследования. 28 (2): 235–244. Дои:10.1016 / j.cretres.2006.07.003. Архивировано из оригинал (PDF) на 2017-10-25. Получено 2017-12-03.
- ^ Evans, S.E .; Wang, Y .; Джонс, M.E.H. (2007). «Совокупность скелетов ящериц из нижнего мела Китая». Senckenbergiana Lethaea. 87 (1): 109–118. Дои:10.1007 / BF03043910.
- ^ Amiot, R .; Ван, X .; Чжоу, З .; Ван, X .; Buffetaut, E .; Lécuyer, C .; Ding, Z .; Флейто, F .; Хибино, Т .; Kusuhashi, N .; Mo, J .; Suteethorn, V .; Wang, Y .; Сюй, X .; Чжан, Ф. (2011). «Изотопы кислорода динозавров Восточной Азии демонстрируют исключительно холодный раннемеловой климат». Труды Национальной академии наук. 108 (13): 5179–5183. Дои:10.1073 / pnas.1011369108. ЧВК 3069172. PMID 21393569.
















