Гигантораптор - Gigantoraptor
| Гигантораптор | |
|---|---|
 | |
| Установлен реконструированный скелет Гигантораптор в Япония | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Надсемейство: | †Caenagnathoidea |
| Семья: | †Caenagnathidae |
| Род: | †Гигантораптор Сюй и другие., 2007 |
| Типовой вид | |
| †Гигантораптор erlianensis Сюй и другие., 2007 | |
Гигантораптор (что означает "гигантский захватчик") является род большого овирапторозавр динозавр который жил в Азия вовремя Поздний мел период. Это известно из Формация Ирен Дабасу из Внутренняя Монголия, где первые останки были найдены в 2005 году.
Гигантораптор достигли огромных размеров; По оценкам, он имеет длину 8–8,9 м (26–29 футов) и весит от 2 до 2,7 т (от 2 000 до 2 700 кг). У него был сильно пневматизированный позвоночник и удлиненные руки с ногами. Обе бедренная кость и большеберцовая кость их длина превышала 1 м (3,3 фута), что необычно среди гигантских теропод. В нижняя челюсть мы беззубый и закончился ороговевшим клюв, как и у других овирапторозавров. Хотя известно, что несколько видов овирапторозавров развили полную шерсть перья, Гиганторапториз-за своего размера мог потерять часть этого покрова.
Род классифицируется как динозавр-овирапторозавр, группа обычно мелких пернатых животных. Хотя первоначально было установлено, что это базальный овирапторид, последующий анализ показал, что это Caenagnathid. Это был гигантский наземный дом двуногий всеядный со срезанным прикусом, на что указывает сохранившаяся нижняя челюсть. Форма клюва указывает на универсальную диету с потенциально редким хищничеством. Голотип - и единственный известный экземпляр - был определен как представляющий молодого взрослого человека, умершего в возрасте 11 лет и достигшего юношеского возраста около 7 лет жизни. Такое развитие указывает на ускоренный рост по сравнению с другими более крупными тероподами. Открытие и исследование яиц крупных овирапторозавров, Macroelongatoolithus, указывает на то, что крупные виды, такие как Гигантораптор строили свои гнезда с отсутствием яиц в центре, чтобы не раздавить.
История открытия
В карьере на Сайхангаоби, Формация Ирен Дабасу, Эрлиан бассейн Сонид Левое знамя (Внутренняя Монголия ), многочисленные остатки зауропод Сонидозавр открыты с 2001 года. Китайский палеонтолог Сюй Син попросили воспроизвести открытие Сонидозавр в апреле 2005 г. для японского документального фильма. Сюй оказал им услугу, выкопав бедренную кость. Вытирая кость, он внезапно понял, что это был не зауропод, а неопознанный теропод размером с Альбертозавр. Затем он прекратил съемку, чтобы обезопасить свою счастливую находку. Таким образом, открытие Гигантораптор голотип ископаемое было задокументировано на пленке.[1][2]
Который голотип, LH V0011, состоит из неполных и диссоциированных останков одного взрослого человека, сохраняющих почти полный нижняя челюсть, частично изолированные шейный позвонок, спинные позвонки, хвостовые позвонки, верно лопатка, верно плечевая кость, верно радиус и локтевая кость, почти полное право манус, частичное подвздошная кость с почти полным лобок и задние конечности, включая обе бедренная кость, большеберцовая кость и малоберцовая кость с очень полным пес.[1]
В 2007 г. типовой вид Гигантораптор erlianensis был назван и описан Сюй, Тан Цинвэй, Ван Цзяньминь, Чжао Сицзинь и Тан Лин. В родовое имя, Гигантораптор, получается из латинский гига, гигантис, смысл "гигант " и раптор, что означает «захватчик». В конкретное имя, Erlianensis, относится к Эрлианский бассейн.[1] Голотип в настоящее время хранится в Институте геологии и палеонтологии Лунхао, Внутренняя Монголия.[3]
Описание
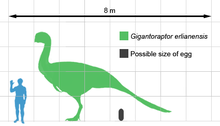
Гигантораптор является самым крупным из известных овирапторозавров, у которых имеется скелетный материал. Примерно в 3 раза длиннее и в 35 раз тяжелее, чем самый крупный ранее обнаруженный овирапторозавр. Читипати, голотип Гигантораптор был оценен в длину 8 м (26 футов), высоту в бедрах 3,5 м (11 футов) и тяжелый вес 1,4 т (1,400 кг).[1] Другие оценки предполагают длину 8,9 м (29 футов) и массу от 2 до 2,7 т (от 2000 до 2700 кг).[4][5][6]
Достаточно известно, что овирапторозавры имеют развитые перья, как это видно на пернатых экземплярах Каудиптерикс и пигостиль нескольких овирапторид.[1][7] Как овирапторозавр, Гигантораптор вероятно, тоже был оперенным, однако, учитывая его огромные размеры, он мог быть частично голым.[1]
Череп



В сохраненном состоянии общая длина нижней челюсти 46 см (460 мм). Этот элемент слился в широкую лопату, что указывало на то, что неизвестный череп был более полуметра в длину и беззубый, вероятно, оснащен рамфотека (роговой клюв). По сравнению с правой стороной, левая сторона челюстей хорошо сохранилась с почти нетронутой формой. Зубные кости очень глубокие и беззубый, а по верхним краям имеют пару резко выраженных полочек. Эти конструкции относительно плоские. Передний конец зубной кости округлый, больше похож на ценагнатиды и отличается от подбородка из некоторых овирапториды. При виде сверху симфизарный (сращение кости) передняя часть зубной кости имеет U-образную форму. Вдоль боковых поверхностей этой кости некоторое количество питательных веществ. отверстие можно найти, который, вероятно, поддерживал рамфотеку.[3]
Обе угловатый и суставно-надуглово-венечный (ASC) комплекс сравнительно меньше зубного. Угловой прочно срастается с комплексом ASC и придает форму нижнему краю внешнего нижнечелюстное отверстие. Угловой имеет изогнутую форму и в основном расширен вперед с большим боковым углублением, которое соединяется с задним нижним процесс (костный выступ / расширение) зубной кости. Комплекс ASC состоит из слияния суставной и надугловой кости с венечный отросток нижней челюсти. Этот комплекс простирается вперед, чтобы соединиться с задним верхним отростком зубной кости и придает форму верхнему краю наружной нижней челюсти. фенестра. Выступ венечного отростка невысокий, отчетливо округлый. На заднем конце нижних челюстей может располагаться суставной гленоид. Он относительно большой, выпуклый и чашеобразный.[3]
Посткраниальный скелет

Передние хвостовые позвонки имеют очень длинные нервные отростки и сильно пневматизированы с глубокими шипами. плевроцели. Средняя часть относительно короткого хвоста несколько усилена длинным презигапофизы. Хвостовые позвонки сильно пневматизированы губчатым костным наполнением. Передние конечности довольно длинные из-за вытянутой формы. плечевая кость (73,5 см (735 мм) в длину) и тонкое манус. Плечевая кость чрезвычайно сильно изогнута наружу и имеет очень округлую голову, как у некоторых птиц. Первый пястный очень короткий, с сильно расходящимся большим пальцем. Задние конечности хорошо развиты и сильно вытянуты: бедро тонкое, с отчетливыми головкой и шеей, размером 1,10 м (110 см); большеберцовая кость имеет одинаковую верхнюю и нижнюю границы и имеет длину 1,10 м (110 см); и плюсна имеет длину 58,3 см (583 мм), причем III плюсневая кость является самой большой. В пес прочный, с большими и сильно изогнутыми ногтями педали.[1][8]
Отличительные характеристики
По словам Сюй и другие. 2007, Гигантораптор можно отличить по следующим признакам: уменьшенная нижняя челюсть, длина которой на 45% меньше длины бедренной кости; передний хвостовые позвонки с удлиненными нервные отростки сзади расположенная коренастая, стержневидная поперечная процессы; зубной с удлиненным задне-вентральным отростком, доходящим до уровня гленоид; редуцированный сзади сужающийся ретро суставной отросток значительно глубже его ширины; передний и средний хвостовые позвонки с большим пневматический отверстие на вентральной поверхности; зубной с двумя ямка на боковой поверхности и близко к наружной нижней челюсти фенестра; передний каудальный центр с задневентральным краем, идущим вентрально; развитая ламинальная система передних хвостовых позвонков; вертикальный презигапофизарный суставные фасетки расположен проксимальнее дистального конца отростка в среднем хвостовом позвонке; лопатка с выраженной выпуклостью вентрально от акромиального отростка на боковой поверхности; уменьшенный пяточная кость скрыто от переднего обзора расширенным астрагал; дистальный предплюсны IV с проксимальным выступом на латеральном крае; передние хвостовые позвонки состоят из опистоцелей, амфикоелей и прокелей; плевроцели присутствует на большинстве хвостовых позвонков; центр переднего хвостового позвонка с парой вертикально расположенных пневматических отверстий на боковой поверхности; поклонился плечевая кость с выступающей сферической вершиной и изогнутым дельтовидно-грудным гребнем; проксимальный отдел плечевой кости с центрально суженным толстым гребнем, идущим по заднему краю; субкруглая вогнутая проксимальная суставная поверхность прямой локтевой кости; плюсневой III с гинглимоидным дистальным концом; суженная проксимальная суставная поверхность и две боковые шлицы на педали ногтевые; радиус с субсферическим дистальным концом; выпуклый медиальный край проксимального конца и медиального мыщелок в три раза удлинен и простирается дальше дистальнее, чем латеральный, на дистальном конце пястный Я; заметный дорсолатеральный отросток на проксимальном конце и продольный шип на вентральном крае проксимальной трети конца второй пястной кости; узкий шлиц-середина к вертельный голова идет вниз до заднего края бедренного конца, а на передней поверхности дистального конца имеется шпонка надколенника; ручные когти с треугольными боковыми шлицами; лобок сжатие с боков; бедро с прямым концом; шейка зажата между ориентированной сзади-медиально сферической вершиной бедренной кости и расширенным переднезадним вертельным гребнем, который более коренастый и выше спереди, чем сзади.[1]
Классификация
В 2007 году Сюй и другие. назначенный Гигантораптор к Oviraptoridae, в базальном положении. Анатомия Гигантораптор включает диагностические функции Овирапторозавры. Однако он также включает в себя несколько функций, обнаруженных в более производных эуманирапторан динозавров, таких как соотношение передних и задних конечностей 60%, отсутствие расширения дистального лопатка и отсутствие четвертый вертел на бедре. Несмотря на размер, Гигантораптор таким образом, был бы более похож на птицу, чем его более мелкие родственники-овирапторозавры.[1]
В 2010 г. был проведен второй анализ Гигантораптор отношения обнаружили, что он был членом Caenagnathidae а не овирапторид.[9] Филогенетический анализ, проведенный Ламанной. и другие. (2014), поддержали это Гигантораптор был базальным ценагнатидом.[10]

В кладограмма ниже следует филогенетический анализ, проведенный Funston & Currie в 2016 г., обнаружил Гигантораптор быть Caenagnathid.[11]
| Caenagnathidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Рост и развитие

А гистологический анализ выполнен на голотипе малоберцовая кость Сюй и его коллеги в 2007 году показали, что образец имел 7 линии задержанного роста (LAG или годичные кольца) в сохраненном виде. Поскольку несколько LAG отсутствовали, команда использовала обратные вычисления для их определения, в результате чего в итоге получилось 11. Количество LAG указывает на то, что голотипу было около 11 лет на момент смерти и он имел средний рост 128–140 кг ( 282–309 фунтов) в год. Проведенная гистология также предполагает, что этот человек был взрослым, поскольку имело место обширное развитие вторичной остеоны, плотно упакованные LAG близко к периферии кости и плохо васкуляризированный слой пластинчатая кость во внешнем круговом слое, что свидетельствует об относительно неаккуратном росте. Однако, учитывая, что группы LAG с четвертого по седьмой были не очень плотно упакованы, Сюй и его коллеги пришли к выводу, что особь достигла юного совершеннолетия в возрасте 7 лет. Более того, было обнаружено, что образец все еще был относительно молодым взрослым и, следовательно, он стал бы больше, когда достиг бы взрослой стадии. Это указывает на то, что Гигантораптор имели скорость роста намного быстрее, чем у большинства крупных нептичьих теропод, таких как тираннозавриды.[1]
Еще одно необычное развитие в Гигантораптор - удлиненные задние конечности. Сюй и его коллеги отметили, что чем больше становится теропод, тем короче и коренастее становятся ноги, чтобы выдерживать увеличивающуюся массу. Это контрастирует Гигантораптор причем бедра и голени превышают 1 м (3,3 фута), комбинация, которая обычно не встречается у тероподов такого размера. Удлинение этих конечностей указывает на то, что это был один из самых беглый - животное, приспособленное к бегу или поддержанию определенной скорости в течение большого количества времени - большие тероподы. Более того, Гигантораптор у него был сильно пневматизированный позвоночный столб, что могло помочь в похудании. Команда также предложила Гигантораптор мог быть «голым», потому что он в 300 раз тяжелее видов, подобных Каудиптерикс, а крупные животные, как правило, больше полагаются на массу для регулирования температуры, что приводит к потере изоляционных покрытий, которыми обладают их более мелкие сородичи. Однако они предположили, что, по крайней мере, перья рук, вероятно, все еще присутствовали на Гигантораптор, поскольку их основные функции, такие как поведение дисплея и накрывая яйца, пока задумчивый, не связаны с регулированием тепла тела.[1]
Кормление

В 2017 году Вайсум Ма и его коллеги повторно исследовали сохранившиеся челюсти из Гигантораптор и обнаружил, что у него самый глубокий клюв среди ценагнатид и относительно другая диета, чем у других овирапторозавров. Например, суставная область челюстей Гигантораптор демонстрируют сходство с современными Tuatara. Суставная область у этой рептилии обеспечивает движение пропалинальной челюсти (движение вперед и назад) во время кормления, и, основываясь на сходстве их челюстей, Ма и его коллеги предположили, что Гигантораптор также могли использовать пропалинальное движение челюстей. Этому движению способствовали острые полки зубной может указывать на режущий укус, способный прорезать растения (и потенциально мясо ). Это сопоставимо с другими ценагнатидами и контрастирует с челюстями овирапторид, чьи челюсти кажутся лучше подходящими для измельчение пищи. Глубина нижней челюсти свидетельствует о наличии большого язык, которые могли бы помочь животному в обработке пищи и, возможно, повысить эффективность кормления.[3]
Поскольку форма и размер трибуна в большом травоядные животные связано с диетическими привычками, сильно U-образная зубная кость Гигантораптор можно предположить, что этот овирапторозавр был очень неселективный питатель, адаптация, которая могла быть полезной в различных условиях окружающей среды формации Ирен Дабасу. Команда основала эту гипотезу на сравнении с несколькими травоядными животными, включая других динозавров. В то время как резко развитые и узкие ростры больше соответствуют выборочной диете, как в случае некоторых цератопсы, широкие и округлые ростры больше связаны с универсальной диетой, что видно по морфологии Гигантораптор, Евоплоцефал и гадрозавриды. Ма и его коллеги также указали, что его большой размер и, следовательно, более высокие потребности в энергии могли повлиять на его диетические привычки, поскольку более крупные животные, как правило, потребляют пищу более низкого качества, поскольку она доступна в больших количествах в окружающей среде и имеет более стабильные поставки. Однако неизвестно, были ли кишка из Гигантораптор был специализирован для обработки этого большего проглатывание. Команда не могла оставить без внимания случайные плотоядный диета, поскольку сильный клюв вместе с движением пропалинальной челюсти, возможно, позволили переработать мясо.[3]
Размножение

Существование гигантских овирапторозавров, таких как Гигантораптор, объясняет несколько более ранних азиатских находок очень больших, до 53 сантиметров в длину, яиц овирапторозавров, отнесенных к виды Macroelongatoolithus Carlylensis. Они были уложены огромными кольцами диаметром три метра. Наличие Macroelongatoolithus в Северной Америке указывает на присутствие там гигантских овирапторозавров, хотя ископаемых останков скелетов обнаружено не было.[12]
В 2018 году Кохей Танака и его команда исследовали яичные кладки многочисленных экземпляров овирапторозавров, включая кладку яиц Macroelongatoolithus, чтобы соотнести гнездо комплектация и размер корпуса до инкубационное поведение. Их результаты показали, что пористость яичной скорлупы указывает на то, что яйца почти наверняка все овирапторозавры были выставлены в гнезде без внешнего покрытия. Хотя большинство гнезд овирапторозавров имеют яйца, расположенные по кругу, морфология гнезда отличается у более мелких и крупных видов тем, что центр гнезда сильно сокращен у первых видов и становится значительно больше у последних. Такая конфигурация гнезда предполагает, что в то время как самые маленькие овирапторозавры, вероятно, сидели прямо на яйцах, большие, Гиганторапторживотное размером скорее всего сидело на участке, лишенном яиц. Танака и его коллеги отметили, что эта адаптация была полезной для предотвращения раздавливания яиц и могла позволить некоторый контакт с телом во время инкубации у этих гигантских овирапторозавров.[13]
Палеосреда

Гигантораптор известно из Формация Ирен Дабасу из Внутренняя Монголия. Это образование датировано 95,8 ± 6,2 года. миллион лет назад на основе U – Pb и стратиграфический анализы.[14] Окружающая среда вокруг этого образования была очень влажной, существовала большая речная долина с заплетенными поймы. А полузасушливый климат также обозначается Caliche осаждение на основе. Формация Ирен Дабасу имела обширные растительность и листва, о чем свидетельствует палеопочва развитие, многочисленные травоядный динозавры останки, обнаруженные как в русле реки, так и в пойменных отложениях. Богатое разнообразие харофиты и остракоды населяли пойменные системы.[15]
Фауна динозавров этой формации включает быстроходных тираннозавроид Алектрозавр, то орнитомимозавр Археорнитомим, то теризинозавриды Эрлиансавр и Неймонгозавр, то зауропод Сонидозавр, и гадрозавроиды Бактрозавр и Гилмореозавр.[16][17] Ирен Дабасу и Баян Шире образования были сопоставлены на основе их сходной летописи окаменелостей позвоночных. Эта корреляция может быть дополнительно подтверждена наличием гиганта, Гиганторапторразмер Caenagnathid в формации Баян Ширех. Однако, учитывая, что этот образец весьма фрагментарен, его отнесение к Гигантораптор не может быть подтверждено. Присутствие Баян Шире Ценагнатид и Гигантораптор во флювиальных отложениях, таких как формации Ирен Дабасу и Баян Ширех, предполагают предпочтение перед мезические среды обитания в гигант виды овирапторозавров, а не ксерические среды обитания.[18]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k Xing, X .; Tan, Q .; Wang, J .; Чжао, X .; Тан, Л. (2007). «Гигантский птицеподобный динозавр из позднего мелового периода Китая». Природа. 447 (7146): 844−847. Bibcode:2007Натура.447..844X. Дои:10.1038 / природа05849. PMID 17565365. Дополнительная информация
- ^ Пикрелл, Н. (2014). Летающие динозавры: как грозные рептилии превратились в птиц. NewSouth Publishing. 256 стр.
- ^ а б c d е Ma, W .; Wang, J .; Pittman, M .; Tan, Q .; Tan., L .; Guo, B .; Сюй, X. (2017). «Функциональная анатомия гигантской беззубой нижней челюсти птицеподобного динозавра: гигантораптор и эволюция челюсти овирапторозавра». Научные отчеты. 7 (1): 16247. Bibcode:2017НатСР ... 716247М. Дои:10.1038 / s41598-017-15709-7. ЧВК 5701234. PMID 29176627.
- ^ Кампионе, Н.Е .; Evans, D.C .; Brown, C.M .; Каррано, М. Т. (2014). «Оценка массы тела двуногих животных, не являющихся птицами, с использованием теоретического преобразования в пропорции стилоподиальных четвероногих». Методы в экологии и эволюции. 5 (9): 913−923. Дои:10.1111 / 2041-210X.12226. HDL:10088/25281.
- ^ Benson, R. B.J .; Кампионе, Н.Е .; Каррано, М. Т .; Mannion, P.D .; Салливан, К .; Upchurch, P .; Эванс, Д. К. (2014). «Темпы эволюции массы тела динозавров указывают на 170 миллионов лет устойчивых экологических инноваций в линии птичьего стебля». PLOS Биология. 12 (6): e1001896. Дои:10.1371 / journal.pbio.1001853. HDL:10088/25994. ЧВК 4011683. PMID 24802911.
- ^ Molina-Pérez, R .; Ларраменди, А. (2016). Records y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. Барселона, Испания: Ларусс. п. 271. ISBN 9788416641154.
- ^ Лица, W. S .; Брух (2014). «Формы и функции хвоста овирапторозавра». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0093.
- ^ Lü, J .; Карри, П. Дж .; Xu, L .; Чжан, X .; Pu, H .; Цзя, С. (2013). "Овирапторидные динозавры размером с курицу из центрального Китая и их онтогенетические последствия". Naturwissenschaften. 100 (2): 165−175. Bibcode:2013NW .... 100..165л. Дои:10.1007 / s00114-012-1007-0. PMID 23314810.
- ^ Longrich, N.R .; Карри, П. Дж .; Чжи-Мин, Д. (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандаху, Внутренняя Монголия». Палеонтология. 53 (5): 945–960. Дои:10.1111 / j.1475-4983.2010.00968.x.
- ^ Lamanna, M. C .; Sues, H.D .; Schachner, E. R .; Лайсон, Т. Р. (2014). "Новый крупнотелый динозавр-теропод-овирапторозавр из позднего мела Западной Северной Америки". PLOS ONE. 10 (4): e0125843. Дои:10.1371 / journal.pone.0092022. ЧВК 3960162. PMID 24647078.
- ^ Funston, G .; Карри, П. Дж. (2016). «Новый ценагнатид (Dinosauria: Oviraptorosauria) из формации Хорсшу-Каньон в Альберте, Канада, и переоценка родственных связей Caenagnathidae». Журнал палеонтологии позвоночных. 36 (4): e1160910. Дои:10.1080/02724634.2016.1160910.
- ^ Саймон, Д. Дж. (2014). "Яйца гигантских динозавров (тероподов) Oogenus Macroelongatoolithus (Elongaroolithidae) из юго-восточного Айдахо: таксономические, палеобиогеографические и репродуктивные последствия" (PDF). Цитировать журнал требует
| журнал =(помощь) - ^ Tanaka, K .; Зеленицкий, Д.К .; Lü, J .; DeBuhr, C.L .; Yi, L .; Jia, S .; Ding, F .; Xia, M .; Liu, D .; Shen, C .; Чен, Р. (2018). «Инкубационное поведение динозавров-овирапторозавров в зависимости от размера тела». Письма о биологии. 14 (5): 20180135. Дои:10.1098 / rsbl.2018.0135. ЧВК 6012691. PMID 29769301.
- ^ Guo, Z. X .; Shi, Y.P .; Ян, Ю. Т .; Jiang, S. Q .; Li, L.B .; Чжао, З. Г. (2018). «Инверсия бассейна Эрлиан (северо-восток Китая) в начале позднего мела: последствия для столкновения Охотоморского блока с Восточной Азией» (PDF). Журнал азиатских наук о Земле. 154: 49−66. Дои:10.1016 / j.jseaes.2017.12.007.
- ^ Van Itterbeeck, J .; Хорн, Д. Дж .; Bultynck, P .; Ванденберге, Н. (2005). «Стратиграфия и палеоокружающая среда верхнемеловой формации Ирен Дабасу, содержащей динозавров, Внутренняя Монголия, Китайская Народная Республика». Меловые исследования. 26 (4): 699−725. Дои:10.1016 / j.cretres.2005.03.004.
- ^ Карри, П. Дж .; Эберт, Д. А. (1993). «Палеонтология, седиментология и палеоэкология формации Ирен Дабасу (верхний мел), Внутренняя Монголия, Китайская Народная Республика». Меловые исследования. 14 (2): 127−144. Дои:10.1006 / cres.1993.1011.
- ^ Xing, H .; Привет.; Li, L .; Си, Д. (2012). «Обзор изучения стратиграфии, седиментологии и палеонтологии формации Ирен Дабасу, Внутренняя Монголия». В Вэй, Д. (ред.). Материалы тринадцатого ежегодного собрания Китайского общества палеонтологии позвоночных (на китайском языке). Пекин: China Ocean Press. С. 1-44.
- ^ Tsuihiji, T .; Watabe, M .; Barsbold, R .; Цогтбаатар, К. (2015). «Гигантский ценагнатид-овирапторозавр (Dinosauria: Theropoda) из верхнего мела пустыни Гоби, Монголия». Меловые исследования. 56: 60−65. Дои:10.1016 / j.cretres.2015.03.007.
внешняя ссылка
 СМИ, связанные с Гигантораптор в Wikimedia Commons
СМИ, связанные с Гигантораптор в Wikimedia Commons Данные, относящиеся к Гигантораптор в Wikispecies
Данные, относящиеся к Гигантораптор в Wikispecies- [email protected]: "Найден гигантский птицеподобный динозавр".
- Проводная наука Ученые обнаружили в Монголии динозавра-гигантораптора весом 3000 фунтов
- Полная реставрация Гигантораптор erlianensis. Кредит: Юлиус Т. Чотони
- Yahoo! Новости: Китай обнаружил новые виды больших, похожих на птиц динозавров
















