Далианзавр - Daliansaurus
| Далианзавр | |
|---|---|
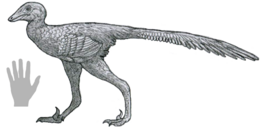 | |
| Реконструкция жизни | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Troodontidae |
| Подсемейство: | †Sinovenatorinae |
| Род: | †Далианзавр Шен и другие., 2017 |
| Типовой вид | |
| Далианзавр liaoningensis Шен и другие., 2017 | |
Далианзавр (смысл "Далянь рептилия ") род малых троодонтид теропод динозавр длиной примерно 1 метр (3 фута 3 дюйма) от Раннемеловой период из Китай. Он содержит один вид, D. liaoningensis, названный в 2017 году Шеном и его коллегами из почти полного скелета, сохранившегося в трех измерениях. Далианзавр необычен тем, что имеет увеличенный коготь на четвертом пальце стопы в дополнение к «серповидному когтю», обнаруженному на втором пальце стопы большинства паравианцы. Он также давно плюсневые кости, и, по-видимому, обладает птичьим крючковые процессы (первый среди троодонтид). В слоях Луцзятунь Формация Исянь, вулканический регион с холодным климатом, Далианзавр жили рядом со своими ближайшими родственниками - Синовенатор, Синусоназус, и Мэй, с которой он образует группу Sinovenatorinae.
Описание
Далианзавр это небольшой, легкий, и, скорее всего, пернатый троодонтид длиной примерно 1 метр (3 фута 3 дюйма),[1] похож по размеру на Синовенатор и Синусоназус.[2] Типичен для взрослых троодонтид,[3][4] то нервные дуги слиты с позвоночник в спинные (туловищные) позвонки единственного известного экземпляра. Общий, Далианзавр очень похож на Синовенатор, Синусоназус, и Мэй, своих ближайших родственников, но тонко от них отличается.[1]
Череп и позвонки
Череп Далианзавр, длина которого составляет 13,8 см (5,4 дюйма), что пропорционально больше, чем у Мэй. Далианзавр также имеет меньшую ноздрю. Проекция предчелюстная кость над ноздрей приплюснутая, а не стержнеобразная в Синовенатор, а другой выступ ниже ноздри прямой, как Синусоназус но в отличие от Синовенатор. Перед глазами есть два отверстия в черепе: предчелюстное отверстие каплевидной формы и большое овальное отверстие. анторбитальное окно. Нравиться Мэй, Синовенатор, и Sinornithoides но в отличие от Синусоназус, нижний край нижней челюсти прямой, а не выпуклый. В верхней челюсти имеется не менее 21 положения зубов, а в нижней - 24; последний меньше чем Синовенатор (27[5]) и другие троодонтиды. Нравиться Синовенатороднако зубы, расположенные ближе к задней части рта, имеют маленькие зазубрины на задних краях. Четыре предчелюстных зуба не загнуты назад, а имеют D-образную форму.[1]
Далианзавр имеет как минимум девять шейные позвонки, вероятно, тринадцать спинных позвонков, пять крестцовые позвонки, и по крайней мере двадцать восемь хвостовых позвонков в хвосте. Из крестцовых позвонков пятый имеет самую широкую поперечные отростки в Далианзавр, а третий - самый широкий в Синовенатор. Передний край отростка также менее вогнут в Далианзавр. В ребра двух спинных позвонков необычно крючковые процессы, в виде изогнутых и стержневидных выступов, аналогичных таковым у Велоцираптор,[6] Линхераптор,[7] и Heyuannia.[8] Эти структуры не имеют аналогов среди троодонтид. Нравиться Sinorthoides, нижний конец передней каудальной шевроны длинные, пластинчатые, направлены назад.[1]
Конечности
В плечевая кость из Далианзавр относительно длинный, с дельтовидно-грудным гребнем, занимающим 24% его длины, аналогично Синовенатор и Sinornithoides. На руке три цифры. Первый пястная кость сильно искривлен в вертикальной плоскости, более чем Sinornithoides; Между тем, вторая пястная кость короче третьей, что верно для обоих Синовенатор и Синусоназус. Чуть ниже первый палец руки самый крепкий, и на когтях этого пальца имеется отчетливо глубокая бороздка на боку. Между тем коготь третьего пальца самый маленький.[1]
Далианзавр имеет подвздошная кость что пропорционально больше по сравнению с бедренная кость чем либо Sinornithoides или же Мэй. Две половины подвздошной кости отчетливо разделены, начиная с уровня третьего крестца, как и Мэй, хотя и на меньшее расстояние. Нижний край задней части подвздошной кости очень вогнутый, а не плоский. Синовенатор. На бедренной кости, которая сильно изогнута, отросток, известный как латеральный гребень, расположен ниже другого отростка, заднего вертела, в то время как они находятся на том же уровне в Sinornithoides. Нравиться Sinornithoides, то большеберцовая кость примерно в 1,4 раза длиннее бедренной кости; это меньше в Синовенатор, в 1,25 раза больше длины бедренной кости.[1]
Пропорционально, Далианзавр намного дольше плюсневые кости чем другие троодонтиды. Нравиться Синовенатор, внешний мыщелок внизу первой плюсневой кости больше внутренней. Вторая плюсневая кость необычно короче четвертой, нижний конец второй заканчивается перед блоком или «суставом» четвертой; противоположное дело в Синовенатор и Синусоназус. Третья плюсневая кость частично закрыта сзади четвертой, в отличие от Tochisaurus; это только частично вызвано дроблением окаменелости.[1]
Среди троодонтид, Далианзавр лучше всего отличается большим когтем на четвертом (крайнем) пальце стопы. В то время как троодонтиды хорошо известны своими серповидными когтями на втором пальце стопы, коготь на четвертом пальце стопы. Далианзавр такой же длины, как и второй, и значительно более прочный. Однако бугорок-сгибатель, мышечное прикрепление, не развит на четвертом копытце.[1]
Открытие и наименование
Далианзавр известен по единственному почти целому скелету, сохранившемуся в трех измерениях. Части образца были реконструированы и не являются подлинным ископаемым материалом, включая заднюю часть черепа и заднюю часть нижней челюсти. Кроме того, на ископаемом отсутствуют плечевые пояса и правая задняя конечность; левая плечевая кость, большая часть таза и несколько последних хвостовых позвонков утоплены в окружающих матрица. Голова и шея ископаемого загнуты назад, а сам хвост загнут вперед; левая рука вытянута, а правая поджата внутрь. Образец найден в Раннемеловой период (~126 Ма Старый,[9] датируется Барремский эпоха) вулканический отложения слоев Луцзятунь Формация Исянь, в селе Луцзятунь возле Бейпяо, Ляонин, Китай. Он хранится в Даляньском музее естественной истории (DNHM) в Далянь, Ляонин, под номером образца ДНХМ D2885.[1]
В 2017 году DNHM D2885 был описан в исследовательской статье, опубликованной в Acta Geologica Sinica и автором Шен Цайчжи, Люй Цзюньчан, Лю Сичжао, Мартин Кундрат, Стивен Брусатт, и Гао Хайлун. Они назвали род Далианзавр в честь города Далянь, где хранится окаменелость, и назвали типовой вид после провинции Ляонин.[1] Позже Шен, Лю, Гао и Кундрат опубликовали гистологический анализ локтевая кость и радиус DNHM D2885, вместе с Масато Хосино и Кентаро Уэсуги, в журнале Историческая биология. Для этого анализа они взяли деструктивную выборку шлифы от середины диафиза костей.[2]
Классификация
А филогенетический анализ был проведен Шеном и его коллегами, которые изменили набор данных Рабочей группы Theropod, чтобы добавить Далианзавр и Синусоназус. Они использовали анализ для выявления черт, которые позволили Далианзавр для помещения в Troodontidae, согласно предыдущим оценкам:[10] зубы многочисленные и плотно прилегающие; перемычка предчелюстной кости, разделяющая ноздри, уплощена; есть бороздка на боковой стороне зубной в нижней челюсти; нервные отростки заменяются неглубокими бороздками ближе к концу хвоста; плюсневые кости асимметричны, причем четвертая более крепкая, чем вторая. Анализ также показал, что Далианзавр сформировал отчетливый эволюционное излучение вместе с другими троодонтидами формации Исянь: Мэй, Синовенатор, и Синусоназус. Эта группа, получившая название Sinovenatorinae, была объединена синапоморфия не наблюдается для Далианзавр, наличие широкой и уплощенной четвертой плюсневой кости.[1]
В пределах Sinovenatorinae, Синовенатор, Синусоназус, и Далианзавр сформировали подгруппу за исключением Мэй. Эту подгруппу различают несколько характеристик, но Шен и его коллеги использовали их для диагностики более широкой Sinovenatorinae: предчелюстная кость составляет всего 10% длины верхней челюсти в нижней части челюсти; есть контакт между предчелюстной костью и носовая кость ниже уровня ноздри; на передних краях некоторых зубов отсутствуют зазубрины; то суставные отростки известные как зигапофизы не срослись в крестцовых отделах; и процесс, известный как противовращающий механизм, расположен позади и над тазобедренным суставом, или вертлужная впадина. Наконец, анализ объединил Далианзавр и Синусоназус за исключением Синовенатор, с двумя разделяющими бифуркацию шевронов на обоих концах около конца хвоста; и слияние больше и меньшие вертлуги на бедренной кости в сплошной вертельный гребень.[1]
Результаты анализа, проведенного Шеном и его коллегами, представлены ниже.[1]
| Дейнонихозаврия |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Длинные плюсневые кости присутствуют в Далианзавр предполагают, что его способности к бегу были развиты лучше, чем у других троодонтид. В то же время возможное наличие крючковидных отростков на его ребрах обеспечило бы усиление грудная клетка, а также повышение эффективности мышц, участвующих в дыхании, как у современных птиц.[1]
Гистология
Тонкие срезы от локтевой кости и лучевой кости голотип образец ДНХМ D2885 свидетельствует о том, что кости лишены вторичных остеоны, или остеоны, образованные ремоделирование костей. Сезонный рост костей очевиден через тонкие срезы; три линии задержанного роста (LAG) разделяют кортикальная кость на четыре отдельные зоны в пределах радиуса, что указывает на то, что человеку было не менее четырех лет, когда он умер. (В пределах локтевой кости есть только два LAG.) Эти оценки считались нижними границами, потому что расширение мозговая полость внутри кости могли стереть дополнительные LAG. Сезонный рост мог быть вызван периодами низких температур; подобное явление наблюдается у современных животных, обитающих в холодном климате, в том числе даже эндотермический млекопитающие.[11] Тем не менее, человек, вероятно, умер в теплый период из-за плотного отложения и присутствия первичных остеонов возле внешней поверхности кости.[2]
По всем четырем зонам сосудисто-нервные каналы в первой зоне более плотные, чем в последующих, что указывает на то, что рост был наиболее интенсивным в течение первого года жизни и быстро уменьшался в дальнейшем. Последующие незначительные колебания скорости роста отражаются чередованием полос остеонов и бессосудистой кости (то есть кости без сосудисто-нервных каналов). Тонкие периферийные полосы роста видны между LAG в двух внешних зонах радиуса, которые составляют внешнюю фундаментальную систему (EFS). Наличие EFS предполагает минимальные темпы роста,[12][13] указывая, что человек, возможно, достиг скелетная зрелость в этом размере. Однако наличие некоторых сосудисто-нервных каналов у поверхности периостальный слой, а также некоторые плохо очерченные остеоны в третьей зоне свидетельствуют о том, что рост еще не завершился на момент смерти. В целом анатомические и гистологические данные свидетельствуют о том, что человек быстро рос, но только в конце малолетний этап жизни. Трехфазная модель роста костей - быстрый рост с последующим замедленным ростом и образованием EFS - также видна в Стенонихозавр.[2][14]
Палеоэкология
В Ляонин регион северо-востока Китая был одним из самых продуктивных ископаемые участки для понимания эволюции пернатых динозавров, включая птиц. Все синовенаторины - Синовенатор, Синусоназус, Мэй, и Далианзавр - жили в пластах Луцзятунь формации Исянь. Вместо строгого сосуществования, возможно, они сменяли друг друга со временем или были географически и экологически изолированы. Шен и его коллеги определили синовенаторины как локализованное излучение троодонтид, обитавших в Ляонине в раннем меловом периоде; подобное излучение произошло со многими микрорапторин дромеозавриды которые жили примерно в одном месте и в одно время. Синевенаторины тоже в мгновение ока сравнивали с дромеозавридами, но таксономически информативные анатомические различия между разными членами группы.[15] Однако возможно, что некоторые из этих различий могут возникать в результате индивидуальных или онтогенетический вариация.[1]
Наряду с синовенаторинами в слоях Луцзятунь обитало множество динозавров. К ним относятся микрорапторин Graciliraptor lujiatunensis; то овирапторозавр Incisivosaurus gauthieri; то орнитомимозавры Шэньчжоузавр восточный[16] и Hexing qingyi;[17] то процедурозаврид тираннозавроид Дилонг парадокс;[16] то титанозавр зауропод Euhelopus sp., основанный на зубах;[18] то орнитопод Jeholosaurus shangyuensis; и цератопсы Пситтакозавр lujiatunensis (включая П. майор и Хуншанозавр хоуи[19]) и Ляоцератопс yanzigouensis. Пситтакозавр lujiatunensis была самой распространенной, известны тысячи экземпляров, за ней следовали троодонтиды и Jeholosaurus shangyuensis. Ископаемые останки птиц из пластов Луцзятунь не известны.[16]
Млекопитающие также присутствовали в слоях Луцзятунь, в том числе Acristatherium yanensis,[20] Gobiconodon zofiae, Juchilestes liaoningensis, Maotherium asiaticus, Meemannodon lujiatunensis, Репеномам гигантский, и R. robustus.[21][22][23] Лягушки Liaobatrachus zhaoi[24] и Мезофрин beipiaoensis[25] также были найдены вместе с ящерицей Далингозавр longidigitus.[26] Многие из окаменелостей Луцзятунь демонстрируют исключительную трехмерную сохранность; Наличие туфогенный песчаник и конгломерат селевой поток отложений предполагает, что животные были убиты внезапно и катастрофически лахар из ближайшего щитовой вулкан.[23] Ископаемые остатки беспозвоночных и растений отсутствуют в слоях Луцзятунь, но пыльца из хвойный растения доминируют в современном[9] Кровати Jianshangou.[27] Среднегодовые температуры воздуха в регионе в барремское и Аптян эпохи, будучи холодными до 10 ° C (50 ° F).[28]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о Shen, C .; Lü, J .; Liu, S .; Kundrát, M .; Brusatte, S.L .; Гао, Х. (2017). «Новый динозавр троодонтид из формации Исянь нижнего мела провинции Ляонин, Китай» (PDF). Acta Geologica Sinica. 91 (3): 763–780. Дои:10.1111/1755-6724.13307.
- ^ а б c d Shen, C .; Lü, J .; Gao, C .; Хосино, М .; Uesugi, K .; Кундрат, М. (2018). «Гистология костей предплечья малого теропод. Далианзавр liaoningensis (Paraves: Troodontidae) из формации Исянь, Ляонин, Китай ". Историческая биология. 31 (2): 253–261. Дои:10.1080/08912963.2017.1360296.
- ^ Russell, D .; Донг, З. (1993). «Почти полный скелет нового динозавра троодонтид из раннего мела бассейна Ордос, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле. 30 (10): 2163–2173. Bibcode:1993CaJES..30.2163R. Дои:10.1139 / e93-187.
- ^ Currie, P.J .; Донг, З. (2001). «Новые сведения о троодонтидах мелового периода (Dinosauria, Theropoda) из Китайской Народной Республики». Канадский журнал наук о Земле. 38 (12): 1753–1766. Дои:10.1139 / e01-065.
- ^ Сюй, X .; Norell, M.A .; Wang, X.-L .; Makovicky, P.J .; Ву, X.-C. (2002). «Базальный троодонтид из раннего мела Китая». Природа. 415 (6873): 780–784. Bibcode:2002Натура.415..780X. Дои:10.1038 / 415780a. PMID 11845206.
- ^ Norell, M.A .; Маковицкий, П.Дж. (1999). «Важные особенности скелета дромеозаврид II: информация из недавно собранных образцов Велоцираптор монгольский". Американский музей Novitates. 3282: 1–45. HDL:2246/3025.
- ^ Сюй, X .; Pittman, M .; Салливан, К .; Choiniere, J.N .; Tan, Q.W .; Clark, J.M .; Norell, M.A .; Шуо, В. (2015). "Таксономический статус дромеозаврид позднего мела Linheraptor exquisitus и его значение для систематики дромеозавров ». Позвоночные PalAsiatica. 53 (1): 29–62.
- ^ Лю, Дж. (2003). «Новый овирапторозаврид (Theropoda: Oviraptorosauria) из позднего мела на юге Китая». Журнал палеонтологии позвоночных. 22 (4): 871–875. Дои:10.1671 / 0272-4634 (2002) 022 [0871: ANOTOF] 2.0.CO; 2.
- ^ а б Chang, S.-C .; Gao, K.-Q .; Чжоу, З.-Ф .; Журдан, Ф. (2017). «Новые хроностратиграфические ограничения формации Исянь с последствиями для биоты Джехол». Палеогеография, палеоклиматология, палеоэкология. 487: 399–406. Bibcode:2017ППП ... 487..399С. Дои:10.1016 / j.palaeo.2017.09.026.
- ^ Makovicky, P.J .; Норелл, М.А. (2004). "Troodontidae". In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 184–195.
- ^ Klevezal, G.A .; Клейненберг, С. (1969). Определение возраста млечных по слоистым структурам зубов и кости [Определение возраста млекопитающих по годичным слоям в зубах и костях] (на русском). Перевод Залкинда, Ф. Иерусалим: Израильская программа научных переводов Press. п. 26.
- ^ Woodward, H.N .; Хорнер, J.R .; Фарлоу, Дж. (2011). «Остеогистологические свидетельства определенного роста американского аллигатора». Журнал герпетологии. 45 (3): 339–342. Дои:10.1670/10-274.1.
- ^ de Andrade, R.C.L.P .; Bantim, R.A.M .; de Lima, F.J .; дос Сантос Кампос, Л .; de Sousa Eleutério, L.H .; Sayão, J.M. (2015). «Новые данные о наличии и отсутствии внешней фундаментальной системы у архозавров». Caderno de Cultura e Ciência, Ano X. 14 (1). Дои:10.14295 / cad.cult.cienc.v14i1.932. ISSN 1980-5861.
- ^ Варриккио, Д.Дж. (1993). «Микроструктура костей тероподового динозавра верхнего мела. Troodon formosus". Журнал палеонтологии позвоночных. 13 (1): 99–104. Дои:10.1080/02724634.1993.10011490.
- ^ Сюй, X. (2002). Окаменелости дейнонихозавров из группы Джехол Западного Ляонина и целурозавровская эволюция (Кандидатская диссертация). Пекин: Высшая школа Китайской академии наук. С. 1–322.
- ^ а б c Сюй, X .; Норелл, М.А. (2006). «Ископаемые останки динозавров, не относящихся к птицам, из нижнемеловой группы Джехол в западном Ляонине, Китай» (PDF). Геологический журнал. 41 (3–4): 419–437. Дои:10.1002 / gj.1044. Архивировано из оригинал (PDF) на 2016-03-03. Получено 2017-10-24.
- ^ Jin, L .; Jun, C .; Годфройт, П. (2012). «Новый базальный орнитомимозавр (Dinosauria: Theropoda) из раннемеловой формации Исянь, Северо-Восточный Китай» (PDF). В Godefroit, P. (ed.). Динозавры Берниссара и наземные экосистемы раннего мела. Жизнь прошлого. Блумингтон: Издательство Индианского университета. С. 467–487. ISBN 978-0-253-35721-2.
- ^ Barrett, P.M .; Ван, X.-L. (2007). "Базальные зубы титанозавров (Dinosauria, Sauropoda) из нижнемеловой формации Исянь провинции Ляонин, Китай". Палеомир. 16 (4): 265–271. Дои:10.1016 / j.palwor.2007.07.001.
- ^ Hedrick, B.P .; Додсон, П. (2013). «Пситтакозавриды Луцзятун: понимание индивидуальных и тафономических вариаций с использованием трехмерной геометрической морфометрии». PLoS ONE. 8 (8): e69265. Bibcode:2013PLoSO ... 869265H. Дои:10.1371 / journal.pone.0069265. ЧВК 3739782. PMID 23950887.
- ^ Hu, Y .; Meng, J .; Li, C .; Ван, Ю. (2010). «Новое базальное эвтериальное млекопитающее из раннемеловой биоты Джехол, Ляонин, Китай». Труды Королевского общества B. 277 (1679): 229–236. Дои:10.1098 / rspb.2009.0203. ЧВК 2842663. PMID 19419990.
- ^ Wang, Y.-Q .; Ху, Ю.-М .; Лизать. (2006). «Обзор последних достижений в изучении мезозойских млекопитающих в Китае» (PDF). Позвоночные PalAsiatica. 44 (2): 193–204.
- ^ Лопатин, А .; Аверьянов, А. (2015). "Гобиконодон (Mammalia) из раннего мела Монголии и ревизия Gobiconodontidae ». Журнал эволюции млекопитающих. 22 (1): 17–43. Дои:10.1007 / s10914-014-9267-4.
- ^ а б Jiang, B .; Fürsich, F.T .; Sha, J .; Ван, Б .; Ню, Ю. (2011). «Раннемеловой вулканизм и его влияние на сохранение окаменелостей в Западном Ляонине, северо-восток Китая». Палеогеография, палеоклиматология, палеоэкология. 302 (3): 255–269. Дои:10.1016 / j.palaeo.2011.01.016.
- ^ Dong, L .; Roček, Z .; Wang, Y .; Джонс, M.E.H. (2013). «Незубые животные из нижнемеловой группы Джехол в Западном Ляонине, Китай». PLoS ONE. 8 (7): e69723. Дои:10.1371 / journal.pone.0069723. ЧВК 3724893. PMID 23922783.
- ^ Wang, Y .; Jones, M.E.H .; Эванс, С. (2007). «Молодой ануран из формации Цзюфотан нижнего мела, Ляонин, Китай» (PDF). Меловые исследования. 28 (2): 235–244. Дои:10.1016 / j.cretres.2006.07.003. Архивировано из оригинал (PDF) на 2017-10-25. Получено 2017-10-24.
- ^ Evans, S.E .; Wang, Y .; Джонс, M.E.H. (2007). «Совокупность скелетов ящериц из нижнего мела Китая». Senckenbergiana Lethaea. 87 (1): 109–118. Дои:10.1007 / BF03043910.
- ^ Chen, P .; Wang, Q .; Zhang, H .; Cao, M .; Li, W .; Wu, S .; Шен, Ю. (2005). «Слой Цзяньшангоу формации Исянь в Западном Ляонине, Китай». Наука в Китае Серия D: Науки о Земле. 48 (3): 298–312. Дои:10.1360 / 04yd0038.
- ^ Amiot, R .; Ван, X .; Чжоу, З .; Ван, X .; Buffetaut, E .; Lécuyer, C .; Ding, Z .; Флейто, F .; Хибино, Т .; Kusuhashi, N .; Mo, J .; Suteethorn, V .; Wang, Y .; Сюй, X .; Чжан, Ф. (2011). «Изотопы кислорода динозавров Восточной Азии демонстрируют исключительно холодный раннемеловой климат». Труды Национальной академии наук. 108 (13): 5179–5183. Дои:10.1073 / pnas.1011369108. ЧВК 3069172. PMID 21393569.
















