Ахиллобатор - Achillobator
| Ахиллобатор | |
|---|---|
 | |
| Диаграмма скелета, показывающая размер и сохранившиеся элементы голотипа | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Dromaeosauridae |
| Clade: | †Эвдромеозаврия |
| Подсемейство: | †Dromaeosaurinae |
| Род: | †Ахиллобатор Perle и другие. 1999 |
| Типовой вид | |
| †Ахиллобатор гигантский Perle и другие. 1999 | |
Ахиллобатор (/əˌkɪлəˈбeɪтɔːr/ ə-КИЛ-ə-ЗАЛИВ-тор; означает «герой Ахилла») - это род больших дромеозаврид динозавр который жил в Азия вовремя Поздний мел период от 96 миллионов до 89 миллионов лет назад на территории современной Формация Баян Шире. Род в настоящее время монотипный, включая только типовые виды А. гигантский. Первые останки были найдены в 1989 году во время монголо-российской полевой экспедиции в г. Монголия и позже описан в 1999 году. С момента первого открытия и описания больше не было найдено ни одного экземпляра. Он представляет собой первого и самого крупного дромеозаврида, известного из формации Баян Ширех.
Это было большое, массивное наземное жилище, двуногий плотоядное животное это был бы активный пернатый хищник охота с увеличенным серпом коготь каждую секунду палец. Предполагаемая длина 5 м (16 футов) при весе около 348 кг (767 фунтов), Ахиллобатор считается одним из крупнейших дромеозавров, наряду с Австрораптор, Дакотараптор и Ютараптор. Наиболее характерная черта Ахиллобатор это примитивный таз с вертикально ориентированной лобковой частью, которая отличается от остальных дромеозаврид. Большинство сохранившихся элементов очень прочны, что необычно для дромеозавров, которые обычно были животными легкого телосложения.
Ахиллобатор классифицируется как таксон дромеозаврид, более конкретно в пределах Эвдромеозаврия, группа гиперплотных дромеозавридов, которые были в основном наземными, а не древесными или амфибиями. В большинстве кладистических анализов Ахиллобатор восстановлен как близкий родственник Дромеозавр и Ютараптор, хотя его часто считают сестринским таксоном последнего. Соотношение коренастых и коротких задних конечностей Ахиллобатор указывает, что это не было беглый - животное, приспособленное к скорости или поддержанию высоких скоростей - более того, прочная морфология верхней челюсти предполагает хищное поведение, основанное на крупной добыче.
История открытия

В 1989 г. во время полевых исследований, проведенных Монгольской и Российской палеонтологической экспедицией в г. пустыня Гоби При исследовании обнажений в Хонгиле, Южно-Центральная Монголия, было сделано много открытий окаменелостей динозавров. Примерно в 5,6 км от населенного пункта Хонгил, большая и в основном разрозненная теропод скелет обнаружен в мелкозернистой, средней песчаник /серый аргиллит который был депонирован еще в Поздний мел эпоха на местности Бурхант, Формация Баян Шире. Сохранившийся экземпляр обнаружен вместе с левым верхняя челюсть сохранение девяти зубы и два альвеолы, четыре шейные позвонки, три спинные позвонки и восемь хвостовые позвонки, почти полный тазовый пояс компрометируя оба лобковые волосы, верно Illium и правильно седалищная кость, обе бедра и влево большеберцовая кость, оставили плюсневые кости III и IV, ручной и педальный фаланги с некоторыми ногтевые, верно скапулокоракоид, изолированные радиус, два ребра и каудальный шевроны. Следы хвоста сухожилия были также обнаружены, но в основном изолированы. Было собрано и исключительно готовый помощником палеонтолога Намсараи Батулсина и хранится как МНУФР-15. Десять лет спустя необычный экземпляр был официально описан в 1999 и стал голотипом для нового рода и вида: Ахиллобатор гигантский. Он был идентифицирован как дромеозаврид таксон, крупный. Описание выполнил монгольский палеонтолог. Алтангерел Перле, и палеонтологи Северной Америки Марк А. Норелл и Джеймс М. Кларк. С точки зрения этимология, то родовое имя, Ахиллобатор, получается из латинский слово Ахиллис (родительный падеж единственное из Ахиллес ) применительно к большому пяточное сухожилие который поддерживал вторую ноготь педали (известный как «серповидный» коготь) большинства дромеозаврид, а старые Монгольский слово баатар (баатар, что означает герой). В конкретное имя, гигантик, получается из Древнегреческий слово γιγαντικός (gigantikós, что означает гигантский) в связи с большим размером голотипа, который превосходит большинство дромеозаврид.[1]
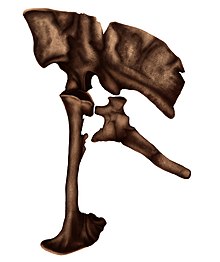
Однако описание было опубликовано в очень предварительном формате, оно было неполным, имело несколько проблем с сохранившимися элементами и многочисленные опечатки.[1] Из-за неправильного толкования, коготь педали II (или серповидный коготь) был заявлен как сохраненный и сочлененный с фалангой педали II, однако это было исправлено Senter в 2007 году, и этот ноготь фактически является ручным.[2] Согласно Тернеру и его коллегам в 2012 году во время обширного пересмотра Dromaeosauridae, статья, вероятно, была опубликована без ведома двух последних палеонтологов, о чем свидетельствует черновик, оставленный в Монголии в 1997 году.[3]
Гипотеза химеры
Тазовый пояс Ахиллобатор Особенности плезиоморфный (примитивный) заурисхий характеристики по сравнению с другими дромеозавридами. Например, лобковая кость выровнена по вертикали и имеет относительно большой лобковый ботинок (широкое расширение на конце), в отличие от большинства других дромеозаврид, у которых, как правило, имеется гораздо меньший ботинок. Сохранившиеся позвонки очень крепкие и имеют ряд плевроцелей.[1] Вышеуказанные различия привели Бернхема и его команду к 2000 предположить, что голотип Ахиллобатор по сути, представляет собой палеонтологический химера, и только ногтевые ножки могли быть получены от динозавров дромеозавридов.[4] Однако на самом деле образец был найден в полусартикулированном виде, и поэтому отнесение останков к одному человеку подтверждено.[5][3] Несмотря на то что Ахиллобатор обладает необычными и примитивными характеристиками по сравнению с другими родственниками, род продолжает восстанавливаться как таксон дромеозавров.[6][7]
Описание

По оценкам, голотип достигал в длину 5 м (16 футов) и весил 348 кг (767 фунтов).[8][9] В большой таксономической ревизии Dromaeosauridae, выполненной Тернером и его коллегами в 2012, они отметили, что этот род представляет второй по величине таксон дромеозаврид (после Ютараптор ) с длиной большеберцовой кости 49 см (490 мм).[3] Гигантский размер Ахиллобатор только приближается или превосходит также паравианцы Австрораптор, Дакотараптор и Ютараптор.[10] Ахиллобатор Это был дромеозаврид с глубоким телом и относительно короткими руками с коренастыми и крепкими задними конечностями. Некоторые из наиболее примечательных особенностей заключались в сильно вертикальном лобке и прочном скелете, таких как глубокая верхняя челюсть и бедро.[1] На голотипе не было следов пух Перо покровов, однако, есть убедительные доказательства, поступающие от других родственников, которые предполагают скорее всего присутствие оперение на Ахиллобатор.[11][12]
Согласно пересмотренному диагнозу Тернера и его коллег в 2012 году, Ахиллобатор можно различить по следующей комбинации характеристик и аутапоморфий: переднечелюстное отверстие полностью обнажено, переднечелюстное и верхнечелюстное отверстия удлиненные и вертикально ориентированные на одном уровне в верхняя челюсть, плюсневой III широкая на верхнем конце, бедренная кость длиннее, чем большеберцовая кость, таз проплобковый, запирательный отросток на седалищной кости большой, треугольной формы расположен на верхней половине седалищного вала, ботинок в дистальном отделе. симфиз лобка развита в краниальном и каудальном аспектах.[3]
Череп
Сохранившиеся верхняя челюсть частично полностью, отсутствует большая часть носового процессы (костлявые выступы). Его длина составляет 29,08 см (290,8 мм), а боковая сторона гладкая по сравнению со спинной частью, хотя его структура очень прочная. В нем сохранено 11 альвеол, из которых 9 заполнены хорошо сохранившимися зубами. Отображение зубов помечено гомодонтия (зубы одинакового размера), с зазубринами и загнутыми назад зубцы немного больше передних зубцов. Наряду с задними зубцами имеется 15 или 18 зубчиков на 5 мм (0,50 см) около центра зубов., Хотя передние зубцы имеют 17-20 зубцов на 5 мм (0,50 см). Учитывая размеры сохранившейся верхней челюсти, Перле и его коллеги предположили, что Ахиллобатор имел относительно большой череп, конкурирующий с черепом карнозавры.[1]
Посткраниальный скелет

Хорошо сохранившийся материал известен из осевой области, представляющей собой очень крепкие и несколько большие шейные, спинные и хвостовые позвонки. Сохранилось около четырех шейных позвонков. По утверждению авторов, передний ряд шейных позвонков развит под острым углом, а передний ряд артикуляция грань центр развита по задней фасетке. Эти характеристики являются индикаторами S-образной формы шеи, как видно из большинства манирапторанс. Сохранились три спинных кости, по-видимому, два спинных ребра. Спинные кости коренастые и имеют плевроцели на боковых поверхностях у некоторых дорсалов по две. Их центральные части сочленения имеют округлую и вогнутую форму. В целом нервные отростки спинных позвонков сходны с таковыми больших позвонков. ратиты такие как вымершие моа и сохранившиеся эму, страус и казуар. Они имеют выступающие межостистые связки шрамы, которые указывают на крепкую и похожую на спину этих птиц. Было найдено восемь хвостовых костей, один из которых относится к переходной серии (передний), три - к средней области и четыре - к заднему концу. Эти четыре были обнаружены в сочленении. Некоторые следы хвоста сухожилия были найдены единичные, вероятно, это признаки оснащенного хвоста. У всех хвостовых позвонков центр амфипластический (плоский с обоих концов), причем фасетка заднего сочленения более вогнутая, чем передняя. Как видно из переходного каудального отдела, передние нервные отростки удлинены и постепенно исчезают на задних каудальных отделах.[1]
Взяв во внимание плечевой пояс, только право скапулокоракоид и радиус сохранились, последний не описан. Однако радиус был измерен как длина 26 см (260 мм). В задней области акромионный отросток, то лопатка сохраняет небольшой бугорок, который прикрепляет область для мышца лопаточно-плечевой в жизни. В гленоид находится в нижней части, обращенной к боковой стороне. Лопатка относительно удлиненная и уплощенная на лопатке. Лезвие утолщено и слегка изогнуто к внутренней стороне, повторяя округлую форму лезвия. грудная клетка. В коракоидный прочный и широкий, его длина составляет 16,7 см (167 мм), а высота - 14,8 см (148 мм). Передний край грубо текстурирован, вероятно, для сочленения с хрящевая ткань. Большой отверстие расположен спереди назад, вероятно, действуя как канал для кровеносный сосуд и нервы.[1]

Обе бедра хорошо сохранились. В целом бедренная кость очень крепкая и длиннее большеберцовой кости, что редко встречается у дромеозаврид, ее длина составляет 50,5 см (505 мм). Он сохраняет подробную информацию о прикрепления мышц. На передней внутренней поверхности есть большая рельефная вогнутая поверхность, которая, вероятно, служила вставкой для m. iliofemoralis internus в жизни, и малый вертел сохраняет вставку для m. pubo ischio femoralis по нижнему краю. На верхней боковой поверхности бедренной кости, снизу ориентированной к большой вертел, имеется бугорок, образовавший прикрепление m. iliofemoralis. Прошивки для m. ilio trochantericus и ischio trochantericus расположены на заднебоковой поверхности на проксимальном конце бедренной кости. На внутренней стороне проксимальной трети стержня сохраняется грубая вогнутая область, вероятно, прикрепление для m. pubo ischio femoralis externus-3. Ниже этой области небольшой выпуклый бугорок присутствовал на прошивке для м.б. pubo ischio femoralis-2. Слева большеберцовая кость немного менее крепкая, чем бедренная кость, но более прямая, ее длина составляет 49 см (490 мм). Поверхность центра вала гладкая, без следов мускулов. Спереди и к задней поверхности синемиальный гребень однако есть крупная и микроскульптурная область, которая служила прикреплением для m. подвздошно-большеберцовая мышца и Ambiens. Дополнительный ткань следы в сохранившейся большеберцовой кости располагаются на нижней поверхности и в задней части. Здесь вероятно углубление с несколько шероховатой поверхностью. апоневроз связанных связки к большеберцовой кости и головка малоберцовой кости когда животное было живым. По-видимому, никаких других поверхностей тканей не сохранилось.[1]
Оставили плюсневые кости III и IV коренастые, размером 23,4 см (234 мм) и 20,9 см (209 мм) соответственно, что указывает на то, что длина плюсны была ниже 50% длины большеберцовой кости. Как и у других манирапторанов, плюсневые кости были тесно соединены. Верхний конец III плюсневой кости уплощен из стороны в сторону и не защемлен на верхнем конце, поэтому отсутствует арктометатарсалий условие. Нижний конец IV плюсневой кости имеет сустав, движение которого ограничено одной плоскостью, а латеральный мыщелок короткий.[1]

Самым символическим элементом голотипа является поразительный таз, практически завершенный; это сохраняет право Illium, седалищная кость и оба лобок. Хотя останки завершены, они немного раздавлены и деформированы. Иллиум заметно высокий: 51,3 см (513 мм) в длину и 28,8 см (288 мм) в высоту с преацетабулярной впадиной, расположенной сверху вниз. На нем много следов тканей: при виде изнутри обе m. каудофеморальный сохраняются. На переднем крае есть микростриции это, вероятно, придало форму m. iliofemoralis в быту, позади этой области, следы м. н. ilio tibialis-2 тоже сохранились. Лобок имеет длину 54,8 см (548 мм) и очень прямой, с большим дистальным голенищем; поразительное сходство с Карнозавры. Например, у других дромеозаврид очень опистопубический конфигурация таза, Ахиллобатор однако имеет примитив, пропубический конфигурация. Большой лобковый ботинок сохраняет очень грубые участки на дорсальной поверхности, индикаторы m. pubo ischio femoralis internus ventralis вложение. Его вид сбоку очень плоский и рельефный, с многочисленными микростризами, которые, вероятно, образовали m. pubo ischio femoralis externus. Сохранившаяся седалищная кость короче последних элементов - 37,8 см (378 мм) в длину. На его стволе образуется большой гребень, который расширяется к переднему краю, вероятно, образуя m. приводящая мышца бедра в жизни. Эта гребнеобразная структура более массивна по сравнению с другими дромеозавридами. Сбоку и в стороне от основного корпуса поверхность очень шероховатая, возможно присоединение m. flexor tibialis internus-1 в жизни[1] как видно в других тероподы и крокодилы.[13][14]
Классификация

Ахиллобатор классифицируется в Dromaeosauridae, группа очень похожих на птиц, манирапторан динозавры. В исходном описании Ахиллобатор был помещен как близкий родственник Дромеозавр с неоднозначным положением в семье.[1] В более поздних и основательных исследованиях этот род считается частью Eudromaeosauria, группы дромеозавридов, которые были облигатными наземными и гиперплотоядными животными, более известными как «настоящие дромеозавриды».[15] Они сильно отличаются от других дромеозавров, таких как древесный микрорапторы[16] или амфибия halszkaraptorines.[17]
Эудромаеозаврия впервые была определена как узловая клады Николасом Р. Лонгричем и Филип Дж. Карри в 2009 как наиболее инклюзивная естественная группа, содержащая Дромеозавр, Велоцираптор, Дейноних, и Saurornitholestes, их последний общий предок и все остальные его потомки. Различные «подсемейства» также были переопределены как клады, обычно определяемые как все виды, более близкие к тезке группы, чем к Дромеозавр или любые однофамильцы из других подклассов.[15]

Недавний филогенетический анализ, проведенный Хартманом и его коллегами в 2019, удивительно восстанавливает Ахиллобатор как сестринский таксон из Иксианозавр,[6] не только это, но Карри и Эванс 2020 восстановили положение рода в Velociraptorinae.[7] Хотя некоторые анализы ставят этот род в необычное положение, Ахиллобатор был обнаружен как дромеозаврин, очень связанный с Ютараптор и Дромеозавр в большинстве филогенетических анализов.[18][3][19][17] Еще больше сходства внутри Ахиллобатор и Ютараптор были обнаружены при повторном остеологическом исследовании Ютараптор экземпляры, поддерживающие их близкие отношения.[20]
При описании Halszkaraptor в 2017 и придуманный Halszkaraptorinae были предложены новые сходства с дромеозавридами, чтобы включить это новое подсемейство. Ахиллобатор был восстановлен в базальном положении вне клады, образованной Дромеозавр и Ютараптор.[17] Но в результатах для Eudromaeosauria филогения на основе филогенетический анализ проведено Ясинским и его коллегами в 2020 во время описания Dineobellator, Ахиллобатор снова был восстановлен в традиционном Ютараптор+Ахиллобатор clade, но на этот раз за пределами Dromaeosaurinae.[21]
Оставили кладограмма как в Cau et al. 2017,[17] и правая кладограмма у Jasinski et al. 2020:[21]
|
|
Палеобиология

Строение задних конечностей и тазовой области Ахиллобатор указывает на то, что у животного был массивный бедра и прочные ноги, подходящие для умеренного быстрого бега. Кроме того, выступающий таз Ахиллобатор имеет собственный специализированный бедренный мышечные ретракторы которые, кажется, указывают на сильную способность к прыжкам.[1] В 2016 году Скотт Персонс IV и Карри исследовали пропорции конечностей многочисленных теропод и обнаружили, что компсогнатиды, троодонтиды и тираннозавроиды были бегущими животными со многими таксонами, извлеченными с относительно высокими показателями CLP (Cursorial-limb-ratio), но дромеозавриды вылавливались с низкими показателями CLP. Ахиллобатор был восстановлен с оценкой -5,3, что указывает на то, что, несмотря на наличие мощных ног, он не был адаптирован к поддерживать высокие скорости на продолжительное время.[22]
Manning et al. 2009 проверили функцию серповидного когтя дромеозаврид, проанализировав биомеханику того, как напряжения и деформации будут распределяться по когтям и конечностям, и используя сравнения в пределах кривизны серповидного когтя дромеозаврид на ступне с кривизной у современных птиц и млекопитающих . Они обнаружили, что они идеально подходят для скалолазания и наземного образа жизни.[23] Питер Мацковицкий заявил, что анализ может быть правильным для примитивных дромеозавридов (таких как Микрораптор ), тем не менее, лазающих по деревьям, это не объясняет, почему гигантские животные любят Ахиллобатор или же Ютараптор сохранили серповидные когти, когда они были слишком большими, чтобы лазить по деревьям. Mackovicky предположил, что более крупные дромеозавриды адаптировали коготь для использования исключительно для более агрессивного хищного поведения.[24]
Поразительное сходство ступней и ног дромеозаврид и таковых у Accipitrid хищные птицы, под руководством Фаулера и др. 2011, чтобы предположить, что дромеозавриды охотились таким же образом, как и те хищный птицы. Они обнаружили, что ступни и ноги дромеозавров похожи на орлы и ястребы имея увеличенный второй коготь и подобный диапазон сгибания, но плюсневые кости имеют большее сходство с таковыми из совы. Модель RPR (Raptor Prey Restraint) предполагает, что дромеозавры прыгнули в свои добыча, обездвиживая его массой своего тела, а затем крепко удерживая большими серповидными когтями; после этого дромеозавр начал питаться животным, пока оно было еще живым, и смерть в конечном итоге наступила от потеря крови и органная недостаточность. Руки или "крылья", которые были скорее всего покрыты длинными перьями (на Чжэньюаньлун[12]), возможно, дромеозавр взмахивал руками, чтобы стабилизировать его равновесие, удерживая добычу, наряду с этим, длинный оперенный хвост, вероятно, работал как противовес основному телу. Наконец, морда была бы полезна для прикончания добычи. С помощью этих наблюдений они установили, что дромеозавриды и троодонтиды были ниша перегородка как крупные хищники, так и мелкие хищники соответственно.[25]
Палеосреда

Остатки Ахиллобатор были раскопаны в местности Бурхант на Формация Баян Шире. Эта формация была разделена на верхнюю и нижнюю части, с некоторыми участками, представляющими границу нижнего и верхнего мела.[26][27][28][29] Бурхант, типовая местность Ахиллобатор, был идентифицирован как местонахождение на границе верхнего мела.[30] Анализы на магнитостратиграфия формации указывают на то, что весь Баян Шире находится в пределах Меловой длинный нормальный, который длился только до конца Сантон этап, давая Сеноманский -Сантон возраст.[31] Недавний кальцит U – Pb анализы , выполненные Курудамой и его коллегами в 2020 году, подтвердили возраст формации Баян Ширех от 95,9 ± 6,0 млн до 89,6 ± 4,0 млн лет назад, что также подтверждает сеномано-сантонский возраст.[32] Со временем сильная корреляция с Формация Ирен Дабасу был предложен многими авторами, в основном на основе сходных комплексов ископаемых.[33][34][29] Эту гипотезу можно также подтвердить близким возрастом.[35]
Наличие Caliche, речной и озерный отложения в различных местах указывают на полузасушливый климат с большими водоемами, такими как реки, озера и потоки.[36][37][33][38] Самойлов и Бенджамини считали свиту Баян-Ширех частично озерной. 1996,[38] однако крупномасштабная перекрестная стратификация на различных слоях песчаника в районе Бэйншира является показателем заметных извилины, помимо этого, подразумевается, что наличие крупных речных водосборов дренировало восточную часть пустыня Гоби.[31] В корнеплоды Ботрокариум бычковатый и Nyssoidea mongolica были зарегистрированы из местонахождения Хара Хутул, где также были обнаружены теризинозавры. Энигмозавр и Сегнозавр. Дополнительные данные об этом месте и Бор-Гуве, кажется, отражают большое присутствие покрытосеменные растения через формацию, с многочисленными ископаемое фрукты восстановленные, которые имеют внешнее сходство с современными окра однако таксономическое родство довольно неясно.[26][39]
Современная палеофауна

Палеофауна которые были современниками Ахиллобатор в свиту Баян-Ширех включены средние тероподы Эрликозавр,[40] Гарудимимус,[41] Сегнозавр[40] и возможно Алектрозавр.[42] Тяжелый анкилозавры Таларурус и Цагантегия.[43] Маленький маркоцефалы Амтоцефал[44] и Грацилицератопс.[45] Базальный гадрозавроид Гобихадрос[30] и большой, с длинной шеей зауропод Эркету.[39] Также присутствовали таксоны, не относящиеся к динозаврам, в основном подвергая опасности полуводных и наземных рептилий, таких как черепахи Линдхолмемис,[46] Gobiapalone и "Трионикс ".[47] Крокодиломорфы представлены Паралигатор[48] и сообщенный ихнотаксон это было сделано плаванием.[49] Рыбы также известны по формации. Примитивный акулы Hybodus и остатки остейхтиан рыбы в разных местах - прекрасные представители.[29]
Судя по всему, было перегородка ниши среди видов, например, высокий браузер теризинозавриды Эрликозавр и Сегнозавр,[40] или пастбище Таларурус и браузер Цагантегия.[43] Эркету, длинношейный зауропод из формации, возможно, был самым высоким травоядным животным.[9] В то время как другие дромеозавриды заполнили множество специализированных экологические ниши как стройный unenlagiines, Ахиллобатор сохранили консервативный образ жизни и заполнили нишу крупнотелых хищник часто от средних до крупных жертв.[15][9][50] Пересмотр максиллы в таксонах эвдромеозавров указывает на то, что азиатские и североамериканские эвдромеозавры были разделены морфологией морды и экологическими стратегиями. Верхняя челюсть, по-видимому, является надежным ориентиром при выводе формы предчелюстная кость и в целом морда. Например, у большинства азиатских видов морда удлиненная, основанная на верхней челюсти (такие животные, как Велоцираптор известны по целым черепам), что указывает на выборочное питание, такое как захват маленькой быстрой добычи. Ахиллобатор тем не менее, это исключение для азиатских эвдромеозавров, отличающихся крепкой и глубокой морфологией челюстей, что указывает на сильную связь и родство с североамериканскими членами, у которых также коренастые и глубокие морды. Адаптации Ахиллобатор и североамериканские эвдромеозавры демонстрируют консервативную экологию, в основном основанную на крупной добыче.[50]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л Perle, A .; Норелл, М. А .; Кларк, Дж. М. (1999). «Новый манирапторан Theropod - Achillobator giganticus (Dromaeosauridae) - из верхнего мела Бурханта, Монголия». Вклад кафедры геологии и минералогии Национального музея Монголии (101): 1–105. OCLC 69865262.
- ^ Сентер, П. (2007). «Метод отличия ручных когтей дромеозаврид от когтей педального серпа» (PDF). Вестник музея естественной истории Гумма (11): 1–6. ISSN 1342-4092.
- ^ а б c d е Тернер, A.H .; Makovicky, P.J .; Норелл, М.А. (2012). "Обзор систематики дромеозаврид и паравианской филогении". Бюллетень Американского музея естественной истории. 2012 (371): 1–206. Дои:10.1206/748.1. HDL:2246/6352. S2CID 83572446.
- ^ Burnham, D.A .; Derstler, K.L .; Currie, P.J .; Баккер, Р.Т .; Чжоу, З .; Остром, Дж. (2000). «Замечательный новый птицеподобный динозавр (Theropoda: Maniraptora) из верхнего мела Монтаны». Палеонтологические материалы Канзасского университета. 13: 1–14. Дои:10.17161 / PCNS.1808.3761. HDL:1808/3761. ISSN 1046-8390.
- ^ Norell, M.A .; Маковицки, П.Дж. (2004). "Dromaeosauridae". In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Калифорнийский университет Press. п. 207. ISBN 9780520941434.
- ^ а б Hartman, S .; Mortimer, M .; Wahl, W.R .; Lomax, D.R .; Lippincott, J .; Лавлейс, Д. (2019). «Новый паравианский динозавр из поздней юры Северной Америки поддерживает позднее приобретение птичьего полета». PeerJ. 7: e7247. Дои:10.7717 / peerj.7247. ЧВК 6626525. PMID 31333906.
- ^ а б Карри, П. Дж .; Эванс, Д. К. (2020). "Черепная анатомия новых образцов Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) из формации динозавров (кампан) в Альберте". Анатомический рекорд. 303 (4): 691–715. Дои:10.1002 / ар.24241. PMID 31497925. S2CID 202002676.
- ^ Тернер, A.H .; Pol, D .; Clarke, J.A .; Эриксон, G.M .; Норелл, М.А. (2007). «Базальный дромеозаврид и эволюция размеров, предшествующая полету птиц». Наука. 317 (5843): 1378–1381. Bibcode:2007Научный ... 317.1378Т. Дои:10.1126 / science.1144066. PMID 17823350. Вспомогательные онлайн-материалы
- ^ а б c Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 151. ISBN 9780691167664.
- ^ Ely, R.C .; Кейс, Дж. А. (2019). «Филогения нового гигантского паравиана (Theropoda; Coelurosauria; Maniraptora) из верхнего мела острова Джеймса Росса в Антарктиде». Меловые исследования. 101: 1−16. Дои:10.1016 / j.cretres.2019.04.003.
- ^ Тернер, A.H .; Makovicky, P.J .; Норелл, М.А. (2007). "Перья гусиные ручки у динозавра Велоцираптора". Наука. 317 (5845): 1721. Bibcode:2007Научный ... 317.1721Т. Дои:10.1126 / science.1145076. PMID 17885130.
- ^ а б Lü, J .; Брусатте, С. (2015). «Большой короткорукий крылатый дромеозаврид (Dinosauria: Theropoda) из раннего мелового периода Китая и его значение для эволюции перьев». Научные отчеты. 11775 (5): 11775. Bibcode:2015НатСР ... 511775Л. Дои:10.1038 / srep11775. ЧВК 4504142. PMID 26181289.
- ^ Ромер, А. (1923). «Тазовая мускулатура заурисхических динозавров» (PDF). Бюллетень Американского музея естественной истории. 48 (19): 605–617.
- ^ Перле, А. (1998). «Примитивные модели структур-мышц тазовых и задних конечностей рептилий (ящериц, крокодилов и птиц)». Монгольский геофизик (9): 26−35.
- ^ а б c Longrich, N.R .; Карри, П.Дж. (2009). «Микрорапторин (Dinosauria – Dromaeosauridae) из позднего мела Северной Америки». Труды Национальной академии наук. 106 (13): 5002–5007. Bibcode:2009ПНАС..106.5002Л. Дои:10.1073 / pnas.0811664106. ЧВК 2664043. PMID 19289829.
- ^ Вешать.; Chiappe, L.M .; Ji, S.-A .; Habib, M .; Тернер, А. Х .; Chinsamy, A .; Лю, X .; Хан, Л. (2014). «Новый динозавр-хищник с исключительно длинным оперением дает представление о летных характеристиках дромеозавров». Nature Communications. 5 (4382): 4382. Bibcode:2014 НатКо ... 5.4382H. Дои:10.1038 / ncomms5382. PMID 25025742.
- ^ а б c d Cau, A .; Бейранд, В .; Voeten, D. F. A. E .; Фернандес, В .; Tafforeau, P .; Stein, K .; Barsbold, R .; Цогтбаатар, К .; Карри, П. Дж .; Годфройт, П. (2017). «Синхротронное сканирование выявляет экоморфологию амфибий в новой стае птицеподобных динозавров». Природа. 552 (7685): 395–399. Bibcode:2017Натура.552..395C. Дои:10.1038 / природа24679. PMID 29211712. S2CID 4471941.
- ^ Senter, P .; Barsbold, R .; Britt, B. B; Бернем, Д. А. (2004). «Систематика и эволюция Dromaeosauridae (Dinosauria, Theropoda)». Вестник музея естественной истории Гунма. 8: 1–20.
- ^ Senter, P .; Kirkland, J. I .; Deblieux, D. D .; Madsen, S .; Тот, Н. (2012). «Новые дромеозавриды (Dinosauria: Theropoda) из нижнего мела штата Юта и эволюция хвоста дромеозаврид». PLOS ONE. 7 (5): e36790. Bibcode:2012PLoSO ... 736790S. Дои:10.1371 / journal.pone.0036790. ЧВК 3352940. PMID 22615813.
- ^ Britt, B.B .; Chure, D. J .; Stadtman, K. L .; Madsen, J. H .; Scheetz, R.D .; Бердж, Д. Л. (2001). «Новые остеологические данные и сходство Ютараптора из горы Кедр (ранний мел) в штате Юта». Журнал палеонтологии позвоночных. 21 (Supp. 3): 36А. Дои:10.1080/02724634.2001.10010852. S2CID 220414868.
- ^ а б Jasinski, S.E .; Sullivan, R.M .; Додсон, П. (2020). «Новый динозавр дромеозаврид (Theropoda, Dromaeosauridae) из Нью-Мексико и биоразнообразие дромеозаврид в конце мелового периода». Научные отчеты. 10 (1): 5105. Bibcode:2020НатСР..10.5105J. Дои:10.1038 / s41598-020-61480-7. ISSN 2045-2322. ЧВК 7099077. PMID 32218481.
- ^ Scott Persons IV, W .; Карри, П. Дж. (2016). «Подход к оценке беглых пропорций конечностей у хищных динозавров и попытка учесть аллометрию». Научные отчеты. 6 (19828): 19828. Bibcode:2016НатСР ... 619828П. Дои:10.1038 / srep19828. ЧВК 4728391. PMID 26813782.
- ^ Manning, P.L .; Margetts, L .; Johnson, M.R .; Холка, P.J .; Продавцы, W.I .; Falkingham, P.L .; Mummery, P.M .; Barrett, P.M .; Раймонт, Д. (2009). «Биомеханика когтей динозавров-дромеозавров: применение рентгеновской микротомографии, наноиндентирования и анализа конечных элементов». Анатомический рекорд. 292 (9): 1397–1405. Дои:10.1002 / ar.20986. PMID 19711472. S2CID 12902686.
- ^ «Смертельные когти Велоцираптора… для лазания». Новый ученый. 203 (2725): 10. 2009. Дои:10.1016 / s0262-4079 (09) 62381-2.
- ^ Fowler, D.W .; Freedman, E.A .; Scannella, J.B .; Камбич, Р. (2011). «Хищная экология дейнонихов и происхождение взмахов крыльев у птиц». PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO ... 628964F. Дои:10.1371 / journal.pone.0028964. ЧВК 3237572. PMID 22194962.
- ^ а б Khand, Y .; Бадамгарав, Д .; Ariunchimeg, Y .; Барсболд Р. (2000). «Меловая система Монголии и ее среды осадконакопления». Меловые среды Азии. Развитие палеонтологии и стратиграфии. 17. С. 49–79. Дои:10.1016 / s0920-5446 (00) 80024-2. ISBN 9780444502766.
- ^ Шувалов, В.Ф. (2003). «Стратиграфия и палеобиогеография мела Монголии». Эпоха динозавров в России и Монголии. Издательство Кембриджского университета. С. 256–278. ISBN 9780521545822.
- ^ Eberth, D.A .; Кобаяши, Ю .; Lee, Y.N .; Mateus, O .; Therrien, F .; Зеленицкий, Д.К .; Норелл, М.А. (2009). «Отнесение Yamaceratops dorngobiensis и связанных с ними красных пластов в Шайн Ус Худаге (восточная часть Гоби, провинция Дорнгоби, Монголия) к переописанной формации Джавхлант (верхний мел)». Журнал палеонтологии позвоночных. 29 (1): 295–302. Дои:10.1080/02724634.2009.10010384. S2CID 197540125.
- ^ а б c Аверьянов А .; Сьюз, Х. (2012). «Соотношение позднемеловых континентальных сообществ позвоночных в Средней и Центральной Азии» (PDF). Журнал стратиграфии. 36 (2): 462–485. S2CID 54210424.
- ^ а б Цогтбаатар, К .; Weishampel, D.B .; Evans, D.C .; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой формации Байншир в пустыне Гоби (Монголия)». PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. Дои:10.1371 / journal.pone.0208480. ЧВК 6469754. PMID 30995236.
- ^ а б Hicks, J.F .; Бринкман, Д.Л .; Nichols, D.J .; Ватабе, М. (1999). «Палеомагнитный и палинологический анализ пластов от альба до сантона в Байн-Шире, Бурхант и Хурен-Дух, восточная часть пустыни Гоби, Монголия». Меловые исследования. 20 (6): 829–850. Дои:10.1006 / cres.1999.0188.
- ^ Kurumada, Y .; Aoki, S .; Aoki, K .; Kato, D .; Saneyoshi, M .; Цогтбаатар, К .; Windley, B.F .; Исигаки, С. (2020). «Кальцитовый U – Pb возраст меловой формации Байн Шире, несущей позвоночных, в пустыне Восточная Гоби в Монголии: полезность каличе для определения возраста». Терра Нова. 32 (4): 246–252. Дои:10.1111 / тер.12456.
- ^ а б Jerzykiewicz, T .; Рассел, Д. А. (1991). «Позднемезозойская стратиграфия и позвоночные животные Гобийской впадины». Меловые исследования. 12 (4): 345−377. Дои:10.1016/0195-6671(91)90015-5.
- ^ Tsuihiji, T .; Watabe, M .; Barsbold, R .; Цогтбаатар, К. (2015). «Гигантский ценагнатид-овирапторозавр (Dinosauria: Theropoda) из верхнего мела пустыни Гоби, Монголия». Меловые исследования. 56: 60−65. Дои:10.1016 / j.cretres.2015.03.007.
- ^ Guo, Z. X .; Shi, Y.P .; Ян, Ю. Т .; Jiang, S. Q .; Li, L.B .; Чжао, З. Г. (2018). «Инверсия бассейна Эрлиан (северо-восток Китая) в начале позднего мела: последствия для столкновения Охотоморского блока с Восточной Азией» (PDF). Журнал азиатских наук о Земле. 154: 49–66. Bibcode:2018JAESc.154 ... 49G. Дои:10.1016 / j.jseaes.2017.12.007.
- ^ Васильев, В.Г .; Волхонин, В.Ц .; Гришин, Г.Л .; Иванов, А.К .; Маринов, И.А .; Мокшанев, К.Б. (1959). Геологическое строение Монгольской Народной Республики (стратиграфия и тектоника). п. 492.
- ^ Мартинсон, Г. (1982). «Верхнемеловые моллюски Монголии». Совместная Советско-Монгольская палеонтологическая экспедиция.. 17: 5–76.
- ^ а б Самойлов, В.С .; Бенджамини, К. (1996). «Геохимические особенности останков динозавров из пустыни Гоби, Южная Монголия». ПАЛАИ. 11 (6): 519–531. Bibcode:1996Палай..11..519С. Дои:10.2307/3515188. JSTOR 3515188.
- ^ а б Ксепка, Д.Т .; Норелл, М.А. (2006). «Эркету эллисони, длинношеий зауропод из Бор Гуве (Дорноговский аймак, Монголия)» (PDF). Американский музей Novitates. 2006 (3508): 1–16. Дои:10.1206 / 0003-0082 (2006) 3508 [1: EEALSF] 2.0.CO; 2.
- ^ а б c Zanno, L.E .; Цогтбаатар, К .; Чинзориг, Т .; Гейтс, Т. (2016). «Специализации анатомии нижней челюсти и зубных рядов Segnosaurus galbinensis (Theropoda: Therizinosauria)». PeerJ. 4: e1885. Дои:10.7717 / peerj.1885. ЧВК 4824891. PMID 27069815.
- ^ Кобаяши, Ю .; Барсболд Р. (2005). «Повторное исследование примитивного орнитомимозавра, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda), из позднего мела Монголии» (PDF). Канадский журнал наук о Земле. 42 (9): 1501−1521. Bibcode:2005CaJES..42.1501K. Дои:10.1139 / e05-044. HDL:2115/14579.
- ^ Перле, А. (1977). "О первой находке Alektrozavra (Tyrannosauridae, Theropoda) из позднего Mela Mongolii" [О первом открытии Alectrosaurus (Tyrannosauridae, Theropoda) в позднем меловом периоде Монголии]. Шинжлех Ухааны Академии геологии в Хуреэлене (на русском). 3 (3): 104–113.
- ^ а б Парк, J.Y .; Lee, Y.N .; Currie, P.J .; Кобаяши, Ю .; Koppelhus, E .; Barsbold, R .; Mateus, O .; Lee, S .; Ким, С. (2020). «Дополнительные черепа Talarurus plicatospineus (Dinosauria: Ankylosauridae) и значение для палеобиогеографии и палеоэкологии бронированных динозавров». Меловые исследования. 108: 104340. Дои:10.1016 / j.cretres.2019.104340.
- ^ Watabe, M .; Цогтбаатар, К .; Салливан, Р. (2011). «Новый пахицефалозаврид из формации Байншир (сеноман-поздний сантон), пустыня Гоби, Монголия» (PDF). Летопись окаменелостей 3. Музей естественной истории и науки Нью-Мексико, Бюллетень. 53: 489–497.
- ^ Серено, П. (2000). «Летопись окаменелостей, систематика и эволюция пахицефалозавров и цератопсов из Азии» (PDF). Эпоха динозавров в России и Монголии. Издательство Кембриджского университета. С. 489–491.
- ^ Суханов, В.Б .; Данилов, И.Г .; Сыромятникова, Е. (2008). «Описание и филогенетическое положение новой черепахи Nanhsiungchelyid из позднего мела Монголии». Acta Palaeontologica Polonica. 53 (4): 601–614. Дои:10.4202 / приложение.2008.0405.
- ^ Данилов, И.Г .; Hirayama, R .; Суханов, В.Б .; Suzuki, S .; Watabe, M .; Витек, Н. (2014). «Меловые черепахи с мягким панцирем (Trionychidae) Монголии: новое разнообразие, записи и пересмотр». Журнал систематической палеонтологии. 12 (7): 799–832. Дои:10.1080/14772019.2013.847870. S2CID 86304259.
- ^ Тернер, А.Х. (2015). "Обзор Shamosuchus и Paralligator (Crocodyliformes, Neosuchia) из мелового периода Азии". PLOS ONE. 10 (2): e0118116. Bibcode:2015PLoSO..1018116T. Дои:10.1371 / journal.pone.0118116. ЧВК 4340866. PMID 25714338.
- ^ Lee, Y.M .; Lee, H.J .; Кобаяши, Ю .; Carabajal, A.P .; Barsbold, R .; Fiorillo, A.R .; Цогтбаатар, К. (2019). «Необычное движение, сохранившееся в пределах крокодиловидной тропы из верхнемеловой формации Баяншири в Монголии, и ее палеобиологические последствия». Палеогеография, палеоклиматология, палеоэкология. 533 (109353): 2. Bibcode:2019ППП ... 533j9239L. Дои:10.1016 / j.palaeo.2019.109239.
- ^ а б Пауэрс, М. А .; Салливан, К .; Карри, П. Дж. (2020). «Пересмотр соотношения предчелюстных и верхнечелюстных признаков у Eudromaeosauria (Dinosauria: Theropoda): расходящиеся тенденции в морфологии морфологии азиатских и североамериканских таксонов». Палеогеография, палеоклиматология, палеоэкология. 547 (109704): 109704. Bibcode:2020ПП ... 547j9704P. Дои:10.1016 / j.palaeo.2020.109704.
внешняя ссылка
 СМИ, связанные с Ахиллобатор в Wikimedia Commons
СМИ, связанные с Ахиллобатор в Wikimedia Commons Данные, относящиеся к Ахиллобатор в Wikispecies
Данные, относящиеся к Ахиллобатор в Wikispecies
















