Эрликозавр - Erlikosaurus
| Эрликозавр | |
|---|---|
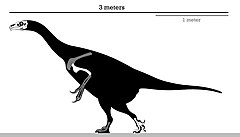 | |
| Схема скелета голотипа | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Надсемейство: | †Теризинозауроидеа |
| Семья: | †Теризинозавры |
| Род: | †Эрликозавр Барсболд & Perle, 1980 |
| Типовой вид | |
| †Эрликозавр andrewsi Барсболд и Перл, 1980 | |
| Синонимы | |
| |
Эрликозавр (что означает «ящерица Эрлика») является род из теризинозаврид который жил в Азия вовремя Поздний мел период. Окаменелости, череп и некоторые посткраниальные фрагменты были найдены в Формация Баян Шире из Монголия в 1972 году, возраст около 96 миллионов 89 миллионов лет назад. Эти останки были позже описаны Алтангерелом Перле и Ринченом Барсболдом в 1980 году, назвав новый род и вид Эрликозавр andrewsi. Он представляет собой второй таксон теризинозавров из этой формации (наряду с Энигмозавр и Сегнозавр ) с наиболее полным черепом среди представителей этого своеобразного семейства динозавров.
В отличие от большинства теризинозаврид, Эрликозавр представлял собой небольшой член, достигавший почти 3,4 м (11 футов) в длину и от 150 до 250 кг (от 330 до 550 фунтов) в массе. У него был хорошо развитый клюв на кончике морды и зубчатые челюсти, которые использовались для его травоядный рацион питания. Ноги закончились на четыре пальцы ног с первым, сочлененным с лодыжка —В отличие от рудиментарный первый палец большинства теропод. Как и другие теризинозавриды, Эрликозавр имел большой кишечник для обработки пищевых продуктов, сильные руки, оканчивающиеся удлиненными когти, и направленный назад таз.
Эрликозавр классифицируется как теризинозавр в пределах Теризинозавры. Теризинозавры были давно загадочными динозаврами с неясными отношениями в первые годы исследований. Последующие исследования подтвердили их истинную природу как динозавров-теропод и систематическое положение среди них. манирапторанс. Клюв и челюсти Эрликозавр указать лист -стригущий способ кормления, характеризующийся активным использованием клюва и шеи. Несколько отличий от сочувствующий Сегнозавр показывает, что эти родственные роды были ниша перегородка.
Открытие и наименование

В голотип образец MPC-D 100/111, найден в слоях из местонахождения Байшин Цав на Формация Баян Шире, состоящий из исключительно хорошо сохранившейся череп, практически полное право пес отсутствует только проксимальный конец плюсны II, III и IV и почти полный левый плечевая кость. Остальные останки включают фрагментарные шейные позвонки Однако количество не указано и они не проиллюстрированы. Эти находки были сделаны во время советско-монгольской экспедиции в г. Омнёгови Провинция в 1972 г.[1][2] Восемь лет спустя род и типовой вид, Эрликозавр andrewsi, был назван и описан (хотя и очень кратко) палеонтологами Ринчен Барсболд и Altangerel Perle в 1980 однако Барсболд не был указан как имяодатель этого конкретного вида. В родовое имя, Эрликозавр, был взят от короля демонов Эрлик, с тюрко-монгольского мифология и Греческий σαῦρος (sauros, что означает ящерица). В конкретное имя, Эндрюси, в честь американского палеонтолога Рой Чепмен Эндрюс, который был руководителем Американских азиатских экспедиций с 1922 по 1930 год.[1] Судя по всему, в первоначальном описании левая стопа была заявлена как часть голотипа,[1] однако это заявление больше не упоминалось.[2][3][4][5]

Как ни странно, в 1981 Перл снова назвал и описал вид, как если бы он был новым, но на этот раз более подробно и написав родовое название как Латинизированный «Эрликозавр».[2] Сегодня большинство авторов считают, что первоначальное название Эрликозавр, действует. На момент открытия это был единственный известный теризинозавр (тогда его называли сегнозаврами.[6]), для которого был обнаружен полный череп, это помогло пролить свет на загадочную и малоизвестную группу динозавров. Он до сих пор представляет собой наиболее известный череп теризинозавра.[4][5]
В 2010, Грегори С. Пол оспаривал действительность этого таксона, утверждая, что Эрликозавр может быть синонимом Энигмозавр (назван в 1983[7]), так как останки последнего были найдены в том же геологическая формация, и известны только по остаткам таза, тогда как таз в Эрликозавр неизвестно; это сделало бы Энигмозавр а младший синоним из Эрликозавр.[8] Однако, поскольку голотип бедра Энигмозавр не очень походил на образец в Сегнозавр как и следовало ожидать от Сегнозавр-подобные остатки ЭрликозаврПалеонтолог Ринчен Барсболд оспорил предполагаемую синонимию, и есть значительная разница в размерах.[9] Вдобавок к этому остатки Эрликозавр и Энигмозавр известны из верхней и нижней границы соответственно.[3][7][10] Как следствие, Энигмозавр и Эрликозавр обычно считаются отдельными родами.[11]
Описание

Поскольку род известен только по очень фрагментарному материалу, было проблематично определить размер Эрликозавр, тем более что большинство позвоночник голотипа отсутствует. Длина черепа голотипа составляет примерно 25 см (250 мм), что указывает на очень маленькую особь. Общий, Эрликозавр был теризинозаврид небольшого размера, по оценкам, достигал около 3,4 м (11 футов) с более легким телосложением, чем тяжелый Сегнозавр.[12] В 2012 Стефан Лаутеншлагер и его коллеги использовали уравнения, специфичные для теропод, для оценки массы тела Эрликозавр и другие теризинозавры. Однако, поскольку бедренная кость неизвестна, они использовали двумерный регрессионный анализ на логарифмически преобразованных данных для Эрликозавр. В результате длина бедренной кости составила 44,33 см (443,3 мм), а вес - 173,7 кг (383 фунта). Учитывая неопределенность этих оценок, они установили общий диапазон масс от 150 до 250 кг (от 330 до 550 фунтов).[5] Согласно альтернативным оценкам, максимальная длина составляет 6 м (20 футов).[9] и более консервативная длина 4,5 метра и вес 500 кг (1100 фунтов).[13] Хотя Эрликозавр в основном не имеет останков тела, как у теризинозаврида, у него было бы сильное сложение рук с большими когтями, широким и громоздким торсом и опистопубическим (направленным назад) тазом.[14] Известно, что теризинозавры были пернатый животных на основе консервированных перьевые оттиски в образцах Бейпиаозавр и Цзяньчангозавр, так что, это скорее всего который Эрликозавр был также пернатым.[15][16]
Череп

Морда умеренно удлиненная, с предчелюстная кость с удлиненным носовым процессы. Тонкая вертикальная пластинка кость Рострально соединен с медиальным краем предчелюстной кости, что указывает на то, что когда животное было живым, хрящевой внутренняя перегородка присутствовал. В дополнение к этому, у предчелюстной кости есть латеральная и медиальная отверстие которые связаны сложной системой сосудистые каналы, который пронизывает структуру предчелюстной кости и, вероятно, связан с сенсорными ветвями нервно-сосудистой сети и глазной нерв поддержка рамфотека (клюв). В верхняя челюсть имеет треугольную форму и сохраняет 24 альвеолы, то зубы находятся гомодонт с грубым зубцы. В зубной имеет клиновидно-удлиненную форму, сохраняет 31 альвеолу. При виде сверху он имеет U-образную форму и сплюснутый сзади с расширением, лежащим поперек. На латеральной и вентральной поверхностях симфизарной области находится ряд отверстий диаметром от 2 до 5 мм (от 0,20 до 0,50 см). Изолированные отверстия внутри соединены сложным сосудисто-нервным каналом. В восстановленном состоянии череп имеет длину 26 см (260 мм), а нижняя челюсть - около 24 см (240 мм).[17]

Хорошо сохранившийся черепная коробка очень полная, отсутствует только сфенетмоидно-мезетмоидный комплекс, тогда как латеросфеноиды и орбитосфеноиды сохраняются не полностью на медиальном виде. Кости вокруг черепной коробки сильно освеженный, но швы между отдельными элементами не видны внешне, за исключением нескольких участков.[3] Однако эти внутренние швы прослеживаются в CT сканирование и, следовательно, элементы корпуса мозга можно было отличить друг от друга.[17] Восстановленный мозг экземпляра несколько удлинен. Обонятельный аппарат и полушария головного мозга очень печально известны, с обонятельный тракт будучи намного больше, чем настоящий мозг. Полушария головного мозга большие и широкие. На поверхности головного мозга можно обнаружить сложные сосудистые борозды, которые обычно находятся в птицы и млекопитающие, а также другие динозавры. Наконец, мозжечок не особо пресловутый, как предыдущие элементы, он удлиненный и коренастый.[5]
Ороговевший клювы, или рамфотеки, хорошо задокументированы среди различных групп внутри Динозаврия.[17] Орнитомимозавры есть веские доказательства этого.[18][19] Однако это не свидетельствует об отсутствии этой анатомической особенности у других групп. Несколько характеристик указывают на рамфотеку, например, беззубая предчелюстная кость с тонким сужающимся нижним краем, последовательная потеря верхних и зубных зубов, вогнутость нижней челюсти на нижней стороне, смещение нижней поверхности зубной кости и ростральная проекция нижнечелюстного симфиза.[17]
В Эрликозавр, наличие ороговевшего клюва на верхней и верхней челюстях можно предположить по наличию многочисленных сосудисто-нервных отверстий на ростральной и боковой поверхности черепа, кроме того, он имеет все упомянутые выше признаки, однако неясно, как протяженность клюв.[17] Сохранившаяся рамфотека у экземпляров Галлимим и Орнитомим свидетельствует о том, что кератиновая оболочка покрывала предчелюстную кость и перекрывала ее с нижней стороны на несколько миллиметров.[18] У некоторых современных птиц рамфотека обычно ограничивается предчелюстной и верхней челюстями, хотя в некоторых случаях у некоторых птиц она частично покрывает носовой отросток.[20] Видимо, в Эрликозавр рамфотека покрывала носовой отросток предчелюстной кости.[17]
Посткраниальный скелет

Остатки тела Эрликозавр очень редки по сравнению с черепными элементами, состоящими из плечевая кость, правая нога и некоторые шейные позвонки. Конкретные шейки матки не были рассчитаны и подсчитаны, а были кратко описаны. Шейные части плоскостопие (слегка вогнутые с обоих концов) с низким нервные дуги.[1] Они относительно прочные, имеют толстую презигапофизы и большие парапофизы. Кроме того, шейки матки имеют некоторое сходство с шейками матки. Сегнозавр, однако, будучи намного меньше.[2]
Сохраненное право пес практически завершена, отсутствует только проксимальный конец плюсневые кости II, III и IV. Он укорочен по длине, с крепкими плюсневыми костями, которые несут расширенные суставные конечности и образуют некомпактную плюсневую кость. I плюсневая кость самая короткая для сравнения, она имеет длину 7 см (70 мм) и расширяет проксимальный суставная поверхность плюсневой кости. Все остальные плюсневые кости примерно равны по размеру, длина второй плюсневой кости составляет 11 см (110 мм). Педаль цифры очень своеобразны по структуре; первая цифра уменьшена по длине, а все остальные цифры почти равны по длине, однако четвертая цифра очень тонкая по сравнению с другими. В фаланги из трех первых цифр укорочены, прочны и имеют сопоставимую структуру. Вторая и третья фаланги четвертого пальца дискообразные, коренастые. Наконец, ногтевые загнуты назад, исключительно крупные, с боков сильно уплощены.[1][2] Грегори С. Пол предположил, что длинная, стройная когти ступней использовались для механизма самообороны.[13]
Слева плечевая кость - единственный сохранившийся остаток грудной области. Плечевая кость имеет удлиненную форму. эпифизы и относительно большой дельтовидный процесс.[1] Он прочный, его расчетная длина составляет 30 см (300 мм). Имеет уменьшенный вал. Проксимальный конец плечевых костей очень широкий. В головка плечевой кости особенности суставной поверхность выпуклая и широкая, посередине суженная к краям. Видный дельто-грудной имеется гребень, вершина которого расположена на 1/3 длины плечевой кости от проксимального конца. Артикуляция мыщелки для радиус и локтевая кость дифференцируются и разделяются укороченной бороздчатой ямка и в целом они очень уменьшены в размере. Ямка для локтевого отростка умеренно глубокая и широкая. Выражается внутренняя шероховатость головы,[2] как в несвязанном Dromaeosauridae.[21]
Классификация

Эрликозавр был назначен Перлом в Segnosauridae,[1] группа, известная сегодня как Теризинозавры, подтвержденный позже кладистический анализы.[4] Теризинозавры были странной группой тероподы которые ели растения вместо мяса и имели обратную сторону лобок, подобно орнитисхианы. Также, как и у орнитисхий, их челюсти заканчивались широким округлым костяком. клюв полезен для обрезки растений.[9][4]
Отношения теризинозавров были довольно сложными, когда были обнаружены первые члены. Например, первый известный таксон теризинозавров, Теризинозавр, был интерпретирован как представляющий черепаха -подобные животные, которые использовали удлиненные когти для кормления водорослей.[22] Однако в 1970 году Рождественский предложил идею, что теризинозавры (тогда известные как сегнозавры), а не существа, не являющиеся динозаврами, на самом деле были тероподы.[23] Позже, в 1980 году, сегнозавры считались медлительными, полуводный животных, при этом Грегори С. Пол утверждал, что у этих спорных животных не было характеристик теропод, и они были прозауроподы с орнитисхийскими приспособлениями они также разделяли эволюционные отношения.[24] Однако при описании большего количества родов, таких как Алксазавр,[25] Наншиунгозавр,[26] и переописание черепа Эрликозавр, стало подтверждаться больше свидетельств о тероподах.[3] С открытием и описанием пернатого Бейпиаозавр, теризинозавры были полностью признаны тероподами, и их начали реконструировать в точной двуногой позе.[15]
Следовательно, теризинозавры теперь классифицируются как теропод, в пределах Целурозаврия. Линдси Занно была одним из первых авторов, которые подробно исследовали родственные связи и родство теризинозавров. Ее работа была полезна во многих филогенетических анализах.[4] Кладограмма ниже является результатом филогенетический анализ в исполнении Хартмана и другие. 2019 используя данные, предоставленные Zanno в 2010 году. Эрликозавр занимал весьма выгодное положение вне клады, образованной Nothronychus graffami и очень сомнительный "Наншиунгозавр" бохлини:[11]
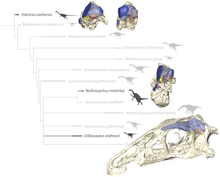
| Теризинозавры |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Чувства

Эрликозавр малоизвестен из посткраниального материала, но голотип черепа стал предметом изучения в Компьютерная томография (КТ) сканы, опубликованные еще в 2012 палеонтологом Стефан Лаутеншлагер и д-р Эмили Рэйфилд из Школы наук о Земле Бристольского университета, профессор Линдси Занно из Музей естественной истории Северной Каролины и Университет штата Северная Каролина и Лоуренс Витмер, профессор палеонтологии Чангского колледжа остеопатической медицины Университета Огайо. Анализ полости мозга показал, что Эрликозаври, вполне вероятно, большинство других теризинозавридов, обладали хорошо развитыми обонянием, слухом и равновесием - чертами, которые лучше ассоциируются с плотоядный тероподы. Увеличенный передний мозг Эрликозавр также мог быть полезен в сложных социальное поведение и уклонение от хищника. Эти чувства были также хорошо развиты у более ранних целурозавров и других теропод, что указывает на то, что теризинозавры, возможно, унаследовали многие из этих черт от своих хищных предков и использовали их для различных и специализированных диетических целей.[5]
В 2019, Грэм М. Хьюз и Джон А. Финарелли проанализировали обонятельная луковица соотношение в современных птицах и сохранившихся черепах нескольких вымерших видов динозавров, чтобы предсказать, сколько гены были вовлечены в обонятельную силу этих вымерших видов. Их анализ показал, что Эрликозавр имело около 477 генов, кодирующих его обонятельные рецепторы, и соотношение обонятельных луковиц 40, что указывает на умеренное обоняние. Оценка Эрликозавр были выше, чем у большинства дромеозавриды несмотря на травоядный образ жизни в этом таксоне, и может отражать переход к сложной социальности и / или снижение зрительных способностей. Хьюз и Финарелли отметили, что по мере того, как динозавры становились больше, размер обонятельной луковицы увеличивался, что может предполагать обоняние как основная сенсорная модальность у крупнотелых нептичьих динозавров.[27]
Кормление и сила укуса

В 2013, Лаутеншлагер выполнил цифровые реконструкции черепной мускулатура в Эрликозавр и обнаружил относительно слабую силу укуса по сравнению с другими тероподами. В целом приводящая мускулатура челюстей, которая в первую очередь предназначена для закрытия челюстей, создает общую силу 374 и 570 Н, но при укусе фактически используется лишь небольшая часть, потому что сила укуса начинает уменьшаться с увеличением расстояния точки укуса приходится на челюстной сустав. Лаутеншлагер обнаружил, что наименьшее усилие на кончике морды составляет 43–65 Н, а наибольшее - в области последнего зуба верхней челюсти - 90–134 Н. Такие факторы, как наличие большого кишечника для обработки растительности и отсутствие повреждений на зубах. зубы предполагают, что Эрликозавр использовал только кончик морды и предчелюстной область, чтобы дотянуться до мягкой листвы или фруктов, и меньшая сила укуса для Эрликозавр лучше подавать в лист -стропильный и растениеводческий механизм кормления, а не активный жевание. В этом исследовании Лаутеншлагер также предположил, что Эрликозавр могли обрабатывать в основном тонкие ветки и растительный материал на основе Стегозавр. Более того, сравнительно узкая ширина морды может указывать на избирательное питание этого теризинозаврида. Наконец, поведение разрыва ветвей Эрликозавр могло быть компенсировано посткраниальной мускулатурой.[28] В том же году Лаутеншлагер и его команда сделали цифровые модели черепа Эрликозавр для проверки функции рамфотека (ороговевший клюв), обнаружив, что эта структура в челюстях действует как структура, снижающая напряжение. Они пришли к выводу, что ороговевшие клювы полезны для повышения устойчивости черепа, делая его менее подверженным изгибам и / или деформации во время кормления.[29]

Хорошо сохранившиеся челюсти также позволили исследованию Бристольского университета определить, как его стиль кормления и диетические предпочтения связаны с тем, насколько широко они могут открывать рот. В исследовании, проведенном Лаутеншлагером и его коллегами в 2015, выяснилось, что Эрликозавр мог открыть рот максимум на 43 градуса. Также в исследование для сравнения были включены плотоядные тероподы. Аллозавр и Тиранозавр. Из сравнений было указано, что у хищных динозавров челюсти были шире, чем у травоядных, как и у современных хищных животных.[30] В 2016, с помощью Метод конечных элементов (МКЭ) и Многотельный анализ динамики (MDA), укус силы Эрликозавр, Платеозавр и Стегозавр были протестированы для оценки пищевых привычек. Результирующая сила укуса для Эрликозавр было между 50-121 N, с черепом, характеризующимся высокой восприимчивостью к нагрузкам и деформации, что указывает на пищевое поведение, специализированное на активном использовании клюва. Результаты дополнительно подтверждают, что Эрликозавр полагался на посткраниальную мускулатуру, чтобы компенсировать низкую силу укуса и снять нагрузку на структуру черепа.[31]
Размножение
Обнаружение окаменелых эмбрионов на Формация Наньчао наличие нескольких яичных гнезд из других формаций указывает на то, что у теризинозавров был колониальный стиль гнездования, подобный другим травоядным, таким как некоторые зауроподоморфы, титанозавры и гадрозавры, поддерживающих социальный образ жизни этих животных. Птенцы теризинозавров были преклонный - способность двигаться после рождения - и не обязательно зависела от родителей.[32] Также известно, что у теризинозавров не было филопатрическое поведение при гнездовании на базе большого гнездового участка в Явхлантская свита, формация, которая покрывает Баян Шире.[33][34]
Палеосреда
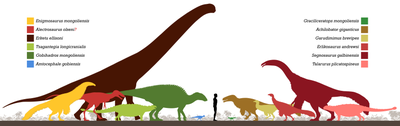
Голотип Эрликозавр был обнаружен в местонахождении Байшин Цав на верхней границе Формация Баян Шире, в карьере, состоящем из серых пески с конгломераты, гравий и серый аргиллиты. Считается, что Байши Цав был сдан на хранение извилистые реки.[35][3][36] Рассмотрение магнитостратиграфия формации, кажется, подтверждает, что весь Баян Шире находится в пределах Меловой длинный нормальный, который длился только до конца Сантонский сцена. Более того, кальцит U – Pb измерения оценивают возраст формации Баян Ширех от 95,9 ± 6,0 млн до 89,6 ± 4,0 млн лет назад, Сеноманский через сантонские времена.[36][37]
Речной, озерный и Caliche седиментация указывает на меньшее полузасушливый климат, при наличии влажных сред, состоящих из больших извилины и озера. Крупномасштабная перекрестная стратификация во многих песчаник слои в местонахождениях Байн Ширех и Буркхант, по-видимому, указывают на большие извилистый реки, и эти большие водоемы могли истощить восточную часть пустыня Гоби.[38][36] Многочисленные окаменелый фрукты были извлечены из населенных пунктов Бор-Гуве и Хара-Хутул.[39] Огромное разнообразие фауна Известно образование, компрометирующее роды динозавров и нединозавров. К товарищам-тероподам относятся большие Ахиллобатор,[40] и дейнохейрид Гарудимимус.[41] Другие травоядные динозавры представлены анкилозавры Таларурус и Цагантегия,[42] маленький маргоцефалы Амтоцефал и Грацилицератопс,[43][44] то гадрозавроид Гобихадрос,[10] и зауропод Эркету.[39] Другая фауна включает полуводный рептилии любят крокодиломорфы и nanhsiungchelyid черепахи.[45][46]
Сосуществование с Сегнозавр

Эрликозавр жили вместе с более крупными видами теризинозаврид в формации Баян Ширех, Сегнозавр. В 2016 году Занно и его коллеги повторно обследовали нижнюю челюсть и зубной ряд из Сегнозавр делая прямые сравнения с таковыми из Эрликозавр в процессе. Они выявили довольно сложные особенности в зубной зубы Сегнозавр, которые представлены наличием многочисленных кили (режущие кромки) и складчатые кили с зубчатыми передними краями, а также увеличение зубчики (зубцы). Вместе эти черты создают шероховатую поверхность у основания зубных коронок, которая была уникальной для Сегнозавр и предположить, что он потреблял уникальные пищевые ресурсы или использовал узкоспециализированные стратегии кормления с добавлением более высокой степени пероральной обработки пищи, чем сочувствующий - родственные виды, которые жили в одном районе в одно и то же время -Эрликозавр. Напротив, у последнего очень симметричные зубы с умеренными зубчиками. Соответствующие нечеткие и специализированные зубные ряды Эрликозавр и Сегнозавр указывает на то, что эти два теризинозаврида были разделены дифференциация ниши в приобретении, переработке продуктов питания или ресурсах. Этот вывод подкрепляется большой разницей в расчетных массах тела, которая составляет до 500%.[47]

В исследовании 2017 года разделения ниш у теризинозавров с помощью цифрового моделирования Лаутеншлагер обнаружил, что более прямые и удлиненные зубные кости примитивных теризинозавров имели самые высокие значения стресса и деформации во время сценариев внешнего питания. В отличие, Эрликозавр и Сегнозавр помогли повернутый вниз кончик нижней челюсти и симфизарный области (сращения костей), а также, вероятно, с помощью клювов, снижающих напряжение и деформацию. Результаты также показали разницу в силе укуса между Сегнозавр и Эрликозавр, что указывает на то, что первый мог бы питаться более жесткой растительностью, в то время как общая устойчивость последнего предполагает большую гибкость в его способе кормления. Лаутеншлагер указал, что эти два таксона были адаптированы к разным способам получения пищи, и что разница в размере и высоте между двумя теризинозавридами еще больше разделяла их ниши. Пока Сегнозавр был адаптирован для использования своих специализированных зубов для заготовки или обработки пищи, Эрликозавр в основном полагался на мускулатуру клюва и шеи при обрезке собирательство.[48]
В дополнение к этим черепным различиям Баттон и Занно в 2019 году отмечают, что травоядные динозавры придерживались двух основных различных режимов питания. Одним из них была обработка пищи в кишечнике, для которой характерны изящные черепа и относительно низкие силы укуса, а вторым - оральная обработка пищи, характеризующаяся особенностями, связанными с обширной обработкой, такой как нижняя челюсть или зубной ряд. Сегнозавр оказался в первом режиме, тогда как Эрликозавр с большей вероятностью попадал во вторую группу, что дополнительно подтверждает, что эти два теризинозаврида были разделены четко определенной нишевой дифференциацией.[49]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм Barsbold, R .; Перле, А. (1980). «Сегнозаврия, новый подотряд хищных динозавров» (PDF). Acta Palaeontologica Polonica. 25 (2): 190−192.
- ^ а б c d е ж Перле, А. (1981). «Новый сегнозаврид из верхнего мела Монголии». Труды совместной советско-монгольской палеонтологической экспедиции (на русском). 15: 50–59. Переведенная статья
- ^ а б c d е Clark, J.M .; Perle, A .; Норелл, М. (1994). «Череп Erlicosaurus andrewsi, позднемелового« сегнозавра »(Theropoda, Therizinosauridae) из Монголии». Американский музей Новитатес. 3115: 1–39. HDL:2246/3712.
- ^ а б c d е Занно, Л. Э. (2010). «Таксономическая и филогенетическая переоценка теризинозаврии (Dinosauria: Maniraptora)». Журнал систематической палеонтологии. 8 (4): 503−543. Дои:10.1080/14772019.2010.488045.
- ^ а б c d е Lautenschlager, S .; Rayfield, E.J .; Perle, A .; Занно, Л. Э .; Витмер, Л. М. (2012). «Эндокраниальная анатомия теризинозаврии и ее значение для сенсорной и когнитивной функции». PLOS ONE. 7 (12): e52289. Bibcode:2012PLoSO ... 752289L. Дои:10.1371 / journal.pone.0052289. ЧВК 3526574. PMID 23284972.
- ^ Перле, А. (1979). «Segnosauridae - новое семейство теропод из позднего мела Монголии» [Segnosauridae - новое семейство теропод из позднего мела Монголии]. Труды совместной советско-монгольской палеонтологической экспедиции (на русском). 8: 45−55. Переведенная статья
- ^ а б Барсболд, Р. (1983). "Хищные динозавры мела Монголии" [Плотоядные динозавры из мелового периода Монголии] (PDF). Труды совместной советско-монгольской палеонтологической экспедиции (на русском). 19: 107. Переведенная статья
- ^ Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. п.159. ISBN 978-0-6911-3720-9.
- ^ а б c Додсон, П. (1993). «Эрликозавр». Эпоха динозавров. Publications International, LTD. п. 142. ISBN 0-7853-0443-6.
- ^ а б Цогтбаатар, К .; Weishampel, D. B .; Evans, D.C .; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой формации Байншир в пустыне Гоби (Монголия)». PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. Дои:10.1371 / journal.pone.0208480. ЧВК 6469754. PMID 30995236.
- ^ а б Hartman, S .; Mortimer, M .; Wahl, W. R .; Lomax, D. R .; Lippincott, J .; Лавлейс, Д. М. (2019). «Новый паравианский динозавр из поздней юры Северной Америки поддерживает позднее приобретение птичьего полета». PeerJ. 7: e7247. Дои:10.7717 / peerj.7247. ЧВК 6626525. PMID 31333906.
- ^ Holtz, T. R .; Рей, Л. В. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов. Случайный дом. ISBN 9780375824197.CS1 maint: дата и год (связь) Список родов для Holtz 2012 Информация о весе
- ^ а б Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. С. 162−168. ISBN 9780691167664.
- ^ Hedrick, B.P .; Занно, Л. Э .; Wolfe, D.G .; Додсон, П. (2015). «Ленивый коготь: остеология и тафономия Nothronychus mckinleyi и N. graffami (Dinosauria: Theropoda) и анатомические особенности производных теризинозаврид». PLOS ONE. 10 (6): e0129449. Bibcode:2015PLoSO..1029449H. Дои:10.1371 / journal.pone.0129449. ЧВК 4465624. PMID 26061728.
- ^ а б Сюй, X .; Tang, Z .; Ван, X.А. (1999). «Теризинозавроидный динозавр с покровными структурами из Китая». Природа. 339 (6734): 350–354. Bibcode:1999Натура.399..350X. Дои:10.1038/20670.
- ^ Pu, H .; Кобаяши, Ю .; Lü, J .; Xu, L .; Wu, Y .; Chang, H .; Zhang, J .; Цзя, С. (2013). "Необычный базальный динозавр теризинозавр с орнитисхийским зубным устройством из Северо-Восточного Китая". PLOS ONE. 8 (5): e63423. Bibcode:2013PLoSO ... 863423P. Дои:10.1371 / journal.pone.0063423. ЧВК 3667168. PMID 23734177.
- ^ а б c d е ж Lautenschlager, S .; Witmer, L.M .; Perle, A .; Занно, Л. Э .; Рэйфилд, Э. Дж. (2014). «Анатомия черепа Erlikosaurus andrewsi (Dinosauria, Therizinosauria): новые идеи, основанные на цифровой реконструкции». Журнал палеонтологии позвоночных. 34 (6): 1263−1291. Дои:10.1080/02724634.2014.874529.
- ^ а б Норелл, М. А .; Makovicky, P.J .; Карри, П. Дж. (2001). «Клюв страусиных динозавров». Природа. 412 (6850): 873–874. Bibcode:2001Натура.412..873Н. Дои:10.1038/35091139. PMID 11528466.
- ^ Барретт П. М. (2005). «Рацион страусиных динозавров». Палеонтология. 48 (2): 347–358. Дои:10.1111 / j.1475-4983.2005.00448.x.
- ^ Кнутсен, Э. М. (2007). Морфология клюва у современных птиц, влияющая на морфологию клюва у орнитомимид (PDF). Университет Осло. п. 44.
- ^ Остром, Дж. Х. (1969). «Остеология Deinonychus antirrhopus, необычного теропод из нижнего мела Монтаны» (PDF). Бюллетень Музея естественной истории Пибоди. 30: 1–165.
- ^ Малеев, Э.А. (1954). "Новый черепахообразный ящер в Монголии". Природа (3): 106–108. Переведенная статья
- ^ Рождественский, А. К. (1970). «На гигантских когтях загадочных мезозойских рептилий». Палеонтологический журнал (1): 131–141.
- ^ Пол, Г.С. (1984). «Сегнозавры динозавры: остатки прозауроподно-орнитисхиевого перехода?». Журнал палеонтологии позвоночных. 4 (4): 507–515. Дои:10.1080/02724634.1984.10012026. ISSN 0272-4634. JSTOR 4523011.
- ^ Рассел, Д. А .; Донг, З. (1993). «Родство нового теропод из пустыни Алкса, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле. 30 (10): 2107–2127. Bibcode:1993CaJES..30.2107R. Дои:10.1139 / e93-183.
- ^ Донг, З. (1979). Динозавры из мелового периода Южного Китая (PDF) (на китайском языке). Мезозойские и кайнозойские красные пласты Южного Китая, Science Press. С. 342–350.
- ^ Hughes, G.M .; Финарелли, Дж. А. (2019). «Размер репертуара обонятельных рецепторов у динозавров». Труды Королевского общества B. 286 (1904): 20190909. Дои:10.1098 / rspb.2019.0909. ЧВК 6571463. PMID 31185870.
- ^ Лаутеншлагер, С. (2013). «Миология черепа и характеристики силы укуса Erlikosaurus andrewsi: новый подход к цифровой реконструкции мышц». Журнал анатомии. 222 (2): 260−272. Дои:10.1111 / joa.12000. ЧВК 3632231. PMID 23061752.
- ^ Lautenschlager, S .; Witmer, L.M .; Perle, A .; Рэйфилд, Э. Дж. (2013). «Эдентулизм, клювы и биомеханические инновации в эволюции тероподных динозавров». PNAS. 110 (51): 20657−20662. Bibcode:2013ПНАС..11020657Л. Дои:10.1073 / pnas.1310711110. ЧВК 3870693. PMID 24297877.
- ^ Лаутеншлагер, С. (2015). «Оценка черепно-мышечно-скелетных ограничений у тероподных динозавров». Королевское общество открытой науки. 2 (11): 150495. Bibcode:2015RSOS .... 250495L. Дои:10.1098 / rsos.150495. ЧВК 4680622. PMID 26716007.
- ^ Lautenschlager, S .; Brassey, C.A .; Баттон, Д. Дж .; Барретт П. М. (2016). «Разъединенная форма и функция в разрозненных кладах травоядных динозавров». Природа. 6 (26495). Bibcode:2016НатСР ... 626495Л. Дои:10.1038 / srep26495. ЧВК 4873811. PMID 27199098.
- ^ Kundrát, M .; Cruickshank, A.R. I .; Manning, T. W .; Nudds, Дж. (2007). «Эмбрионы теризинозавроидных теропод из верхнего мела Китая: диагностика и анализ моделей окостенения». Acta Zoologica. 89 (3): 231−251. Дои:10.1111 / j.1463-6395.2007.00311.x.
- ^ Кобаяши, Ю .; Lee, Y .; Barsbold, R .; Зеленицкий, Д .; Танака, К. (2013). «Первое упоминание о гнездовой колонии динозавров из Монголии показывает гнездовое поведение теризинозавроидов». В Maxwell, E .; Miller-Camp, J .; Анемон Р. (ред.). 73-е ежегодное собрание Общества палеонтологии позвоночных. Лос-Анджелес. п. 155.
- ^ Tanaka, K .; Kobayashi, Y .; Зеленицкий, Д.К .; Therrien, F .; Lee, Y.-N .; Barsbold, R .; Кубота, К .; Ли, Х.-Дж .; Цогтбаатар, Ц .; Идерсайхан, Д. (2019). «Исключительная сохранность места гнездования позднемеловых динозавров из Монголии свидетельствует о колониальном поведении гнездования у нептичьих теропод». Геология. 47 (9): 843−847. Дои:10.1130 / G46328.1.
- ^ Цыбин Ю.И.; Курзанов, С. М. (1979). «Новые данные о верхнемеловых местонахождениях позвоночных Байшин-Цавского района». Труды совместной советско-монгольской палеонтологической экспедиции. 8: 108–112.
- ^ а б c Hicks, J. F .; Бринкман, Д. Л .; Николс, Д. Дж .; Ватабе, М. (1999). «Палеомагнитный и палинологический анализ пластов от альба до сантона в Байн-Шире, Бурхант и Хурен-Дух, восточная часть пустыни Гоби, Монголия». Меловые исследования. 20 (6): 829–850. Дои:10.1006 / cres.1999.0188.
- ^ Kurumada, Y .; Aoki, S .; Aoki, K .; Kato, D .; Saneyoshi, M .; Цогтбаатар, К .; Windley, B.F .; Исигаки, С. (2020). «Кальцитовый U – Pb возраст меловой формации Байн Шире, несущей позвоночных, в пустыне Восточная Гоби в Монголии: полезность каличе для определения возраста». Терра Нова. Дои:10.1111 / тер.12456.
- ^ Самойлов, В. С .; Бенджамини, К. (1996). «Геохимические особенности останков динозавров из пустыни Гоби, Южная Монголия». ПАЛАИ. 11 (6): 519–531. Bibcode:1996Палай..11..519С. Дои:10.2307/3515188. JSTOR 3515188.
- ^ а б Ксепка, Д. Т .; Норелл, М.А. (2006). «Эркету эллисони, длинношеий зауропод из Бор Гуве (Дорногов Аймаг, Монголия)» (PDF). Американский музей Новитатес. 3508 (1): 1–16. Дои:10.1206 / 0003-0082 (2006) 3508 [1: EEALSF] 2.0.CO; 2.
- ^ Perle, A .; Норелл, М. А .; Кларк, Дж. (1999). «Новый манирапторан теропод - Achillobator giganticus (Dromaeosauridae) - из верхнего мела Бурхант, Монголия». Вклад кафедры геологии и минералогии Национального музея Монголии (101): 1–105. OCLC 69865262.
- ^ Кобаяши, Ю .; Барсболд Р. (2005). «Повторное исследование примитивного орнитомимозавра, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda) из позднего мелового периода Монголии». Канадский журнал наук о Земле. 42 (9): 1501–1521. Дои:10.1139 / e05-044. HDL:2115/14579.
- ^ Парк, Дж. (2020). «Дополнительные черепа Talarurus plicatospineus (Dinosauria: Ankylosauridae) и значение для палеобиогеографии и палеоэкологии бронированных динозавров». Меловые исследования. 108: e104340. Дои:10.1016 / j.cretres.2019.104340.
- ^ Серено, П. С. (2000). «Летопись окаменелостей, систематика и эволюция пахицефалозавров и цератопсов из Азии» (PDF). Эпоха динозавров в России и Монголии. Издательство Кембриджского университета. С. 489–491.
- ^ Watabe, M .; Цогтбаатар, К .; Салливан, Р. М. (2011). «Новый пахицефалозаврид из формации Байншир (сеноман-поздний сантон), пустыня Гоби, Монголия» (PDF). Летопись окаменелостей 3. Музей естественной истории и науки Нью-Мексико, Бюллетень. 53: 489–497.
- ^ Данилов, И.Г .; Hirayama, R .; Суханов, В. Б .; Suzuki, S .; Watabe, M .; Витек, Н. С. (2014). «Меловые черепахи с мягким панцирем (Trionychidae) Монголии: новое разнообразие, записи и пересмотр». Журнал систематической палеонтологии. 12 (7): 799−832. Дои:10.1080/14772019.2013.847870. S2CID 86304259.
- ^ Тернер, А. Х. (2015). "Обзор Shamosuchus и Paralligator (Crocodyliformes, Neosuchia) из мелового периода Азии". PLOS ONE. 10 (2): e0118116. Bibcode:2015PLoSO..1018116T. Дои:10.1371 / journal.pone.0118116. ЧВК 4340866. PMID 25714338.
- ^ Занно, Л. Э .; Цогтбаатар, К .; Чинзориг, Т .; Гейтс, Т.А. (2016). «Специализации анатомии нижней челюсти и зубных рядов Segnosaurus galbinensis (Theropoda: Therizinosauria)». PeerJ. 4: e1885. Дои:10.7717 / peerj.1885. ЧВК 4824891. PMID 27069815.
- ^ Лаутеншлагер, С. (2017). «Функциональное разделение ниш у Therizinosauria дает новое понимание эволюции травоядных теропод». Палеонтология. 60 (3): 375−387. Дои:10.1111 / pala.12289.
- ^ Баттон, Д. Дж .; Занно, Л. Э. (2019). «Повторяющаяся эволюция дивергентных видов травоядности у нептичьих динозавров». Текущая биология. 30 (1): 158−168.e4. Дои:10.1016 / j.cub.2019.10.050. PMID 31813611.
внешняя ссылка
- Эрликозавр восстановленный 3D череп в Sketchfab (1)
- Эрликозавр восстановленный 3D череп в Sketchfab (2)
















