Энигмозавр - Enigmosaurus - Wikipedia
| Энигмозавр | |
|---|---|
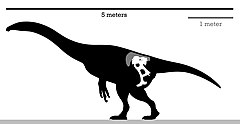 | |
| Схема скелета голотипа | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Clade: | †Теризинозаврия |
| Надсемейство: | †Теризинозауроидеа |
| Род: | †Энигмозавр Барсболд, 1983 г. |
| Типовой вид | |
| †Энигмозавр монгольский Барсболд, 1983 г. | |
Энигмозавр (что означает «Ящерица Энигмы» или «Загадочная ящерица») является род из теризинозавроид который жил в Азия вовремя Поздний мел период. Это было наземное жилище средних размеров, двуногий травоядное животное который представляет третий таксон теризинозавров из Формация Баян Шире, хотя он известен по нижней части. Род монотипный, включая только типовые виды E. mongoliensis, известный по хорошо сохранившемуся тазу и другим предполагаемым останкам тела.
Открытие и наименование

В голотип, IGM 100/84, обнаружен в урочище Хара Хутул в г. Формация Баян Шире (иногда называемая байнширской свитой или Байнширенской свитой), юго-восток Монголия, начиная с Поздний мел период, и впервые было сообщено в 1979 по тазовому сравнению с другими динозаврами теропод. В то время о теризинозаврах было мало что известно.[1] В 1980 году он был снова упомянут, на этот раз в новом инфрапорядке, созданном монгольской палеонтологи Ринчен Барсболд и Алтангерел Перле: Сегнозаврия. По прозвищу «Динозавр из Хара Хутула», он был кратко описан и включен в Сегнозаврию.[2]

Три года спустя типовой вид, Энигмозавр монгольский, был назван и описан в 1983 пользователя Barsbold. Сохранившиеся элементы представляют собой частичный скелет без черепа, включающий хорошо сохранившийся тазовый пояс с другими посткраниями. В родовое имя, Энигмозавр, было заявлено, что происходит от греческого αίνιγμα (аинигма, что означает загадка) и σαῦρος (sauros, что означает ящерица), имея в виду аберратную и необычную форму ее таза. В конкретное имя, mongoliensis, относится к стране его открытия, Монголии.[3] Какой-то теризинозавр ископаемое выводы на Формация Ирен Дабасу сначала считались относящимися к Энигмозавр.[4] Хотя он широко известен по тазу, несколько останков, не упомянутых в первоначальном описании голотипа, были помечены под тем же номером экземпляра (IGM 100/84). Эти элементы находятся в плохом состоянии по сравнению с тазом, и они не были измерены и проиллюстрированы: проксимальный конец бедренная кость; большой диафиз бедренной кости, может быть большеберцовый вал; немного ребра; дистальный конец плечевая кость; предварительный радиус и проксимальный конец локтевая кость. Однако бедренная кость не была обнаружена в связи с голотипом, поэтому ее следует отнести к теризинозаврии. incertae sedis; другие останки могут относиться к голотипу. Очень большое левое бедро длиной 105 см (1050 мм) было помечено тем же номером экземпляра, однако оно не было связано с голотипом из-за его большого размера (больше, чем сам таз). Тем не менее, похоже, он приближается к размеру более близкого Сегнозавр.[5]
Возможная синонимия с Эрликозавр
Немного палеонтологи предложил, чтобы Энигмозавр вероятно было тем же животным, что и Эрликозавр, поскольку оба были найдены в одном геологическая формация, Энигмозавр известен по останкам таза, тогда как таз из Эрликозавр неизвестен и отличительные признаки между ними отсутствовали; если будет доказано, это сделало бы Энигмозавр а младший синоним из Эрликозавр.[6][7] Тем не менее Энигмозавр таз не похож на таз Сегнозавр как и следовало ожидать от связи с Эрликозавр, и разница в размерах огромная.[2][3][8] Кроме того, оба рода известны из разных слоев (нижнего и верхнего),[3][9][10] и большинство авторов считает их отдельными животными.[11][12]
Описание

Энигмозавр был относительно крупным теризинозавром, его длина оценивалась в 5 м (16 футов), а вес составлял от 454 до 907 кг (от 1001 до 2000 фунтов).[13] Как отметил Барсболд в первоначальном описании Энигмозаврего можно узнать по следующим характеристикам: лобковая и седалищная кости короткие; удлиненный край в передней пресимфизарной области дистального отдела лобка.[3] Однако в пересмотренном диагнозе Zanno et al. 2010, есть даже больше характерные черты за Энигмозавр которые не были отмечены / проанализированы ранее: заметные краниальные и каудальные отростки на несколько уплощенном в дорсовентральном направлении лобковом ботинке; лобковые стопы сросшиеся, вытянутые в медиальном направлении, медиальное расширение образует V-образную структуру; медиальное слияние запирательного отростка и лобкового тела образует четырехлучевой отросток.[5]

Таз голотипа относительно хорошо сохранился, с повреждением крестцовых позвонков, частично Illium, Право и лево лобок и слева седалищная кость. Таз в целом несколько большой и опистопубический. Иллиум широко расположен и развернут наружу; в постеродорсальной области сохраняется крупный кубический отросток. Удлиненный лобок цветоносы присутствуют в Иллиуме; однако седалищные ножки более редуцированы. Дистальный конец лобка удлиненный, загнутый и приземистый. Седалищная кость немного короче лобковой кости и параллельна ему, с узким стержнем. Запирательный отросток на переднем крае седалищной кости горизонтально удлиненный и низкий. На большом вертеле заметны открытые края. ямка. Судя по всему, крестец сохраняет шесть позвонков, с удлиненными поперечные отростки.[3] Например, его таз очень специфичен по сравнению с другими родственниками теризинозавров и имеет области резорбция кости и ремоделирование костей на иллиуме. Эти специфические черты могут указывать на преклонный возраст особи, если это правда, слияние запирательного отростка и лобкового тела может быть отклонено как подлинная аутапоморфия для данного вида. Занно отметил, что для решения этой загадки необходимы дополнительные анализы.[5]
Классификация
Энигмозавр был отнесен описателями к отдельной Энигмозавры (ныне устаревший) из-за аномального таза, но позже считался членом Segnosauridae которые сегодня называют Теризинозавры.[3] Линдси Занно в 2010 г. восстановил более базальные позиции в Теризинозауроидеа.[5]

Ниже приведены результаты недавно проведенных филогенетический анализ теризинозаврии Hartman et al. 2019, в котором Энигмозавр восстанавливается как производный теризинозавроид.[12]
| Теризинозаврия |
| |||||||||||||||||||||||||||||||||||||||
Палеоэкология
Остатки Энигмозавр были обнаружены в отложениях, которые были отложены во время Поздний мел период на Формация Баян Шире, Местонахождение Хара Хутул, около 95.9 ± 6.0 млн .-89.6 ± 4.0 млн. Лет назад, Сеноманский -Сантон возрастов.[3][14] Будучи теризинозавром, вероятно, это был медленный травоядное животное и / или всеядный, как постоянно предлагалось большинством авторов.[15][16][17] В биоразнообразие через формацию были характерны теризинозавры, что видно на останках Энигмозавр, Эрликозавр и Сегнозавр.[18] На месте голотипа Хара Хутул также были найдены современные Сегнозавр[2] и безымянный велоцирапторин.[19][20] Большинство оставшихся палеофауна из этой формации известен из верхних слоев, тогда как Энигмозавр известен из нижних слоев.[10]
Здесь обнаружена разнообразная палеофлора: Cornaceae сообщил с Ботрокариум бычковатый и Nyssoidea mongolica как прекрасные представители.[21] Многочисленные ископаемое фрукты находки на местности отражают большое присутствие покрытосеменные растения на формирование. Обнаруженные плоды имеют некоторое сходство с Abelmoschus esculentus однако окончательное таксономическое родство совершенно неясно.[22]
Смотрите также
- Хронология исследований теризинозавров
- Ринчен Барсболд
- Словарь анатомии динозавров
- Анатомические условия расположения
Рекомендации
- ^ Барсболд Р. (1979). «Опистопубический таз у хищных динозавров». Природа. 279 (5716): 792–793. Bibcode:1979Натура 279..792Б. Дои:10.1038 / 279792a0. S2CID 4348297.
- ^ а б c Barsbold, R .; Перле, А. (1980). «Сегнозаврия, новый подотряд хищных динозавров» (PDF). Acta Palaeontologica Polonica. 25 (2): 190–192.
- ^ а б c d е ж грамм Барсболд, Р. (1983). "Хищные динозавры мела Монголии" [Плотоядные динозавры из мелового периода Монголии] (PDF). Труды совместной советско-монгольской палеонтологической экспедиции (на русском). 19: 89. Переведенная статья
- ^ Карри, П. Дж .; Эберт, Д. А. (1993). «Палеонтология, седиментология и палеоэкология формации Ирен Дабасу (верхний мел), Внутренняя Монголия, Китайская Народная Республика». Меловые исследования. 14 (2): 138. Дои:10.1006 / cres.1993.1011.
- ^ а б c d Занно, Л. Э. (2010). «Таксономическая и филогенетическая переоценка теризинозаврии (Dinosauria: Maniraptora)». Журнал систематической палеонтологии. 8 (4): 503–543. Дои:10.1080/14772019.2010.488045. S2CID 53405097.
- ^ Clark, J.M .; Марьянская, Т .; Барсболд Р. (2004). "Теризинозауроида". Динозаврия. Калифорнийский университет Press. п. 159. ISBN 9780520242098.
- ^ Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. п. 159. ISBN 978-0-6911-3720-9.
- ^ Додсон, П. (1993). «Эрликозавр». Эпоха динозавров. Publications International, LTD. п. 142. ISBN 0-7853-0443-6.
- ^ Clark, J.M .; Perle, A .; Норелл, М. (1994). «Череп Erlicosaurus andrewsi, позднемелового« сегнозавра »(Theropoda, Therizinosauridae) из Монголии». Американский музей Novitates. 3115: 1–39. HDL:2246/3712.
- ^ а б Цогтбаатар, К .; Weishampel, D. B .; Evans, D.C .; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой формации Байншир в пустыне Гоби (Монголия)». PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. Дои:10.1371 / journal.pone.0208480. ЧВК 6469754. PMID 30995236.
- ^ Stephan, L .; Emily, J. R .; Perle, A .; Lindsay, E. Z .; Лоуренс, М. В. (2012). «Эндокраниальная анатомия теризинозаврии и ее значение для сенсорной и когнитивной функции». PLOS ONE. 7 (12): e52289. Bibcode:2012PLoSO ... 752289L. Дои:10.1371 / journal.pone.0052289. ЧВК 3526574. PMID 23284972.
- ^ а б Hartman, S .; Mortimer, M .; Wahl, W. R .; Lomax, D. R .; Lippincott, J .; Лавлейс, Д. М. (2019). «Новый паравианский динозавр из поздней юры Северной Америки поддерживает позднее приобретение птичьего полета». PeerJ. 7: e7247. Дои:10.7717 / peerj.7247. ЧВК 6626525. PMID 31333906.
- ^ Holtz, T. R .; Рей, Л. В. (2007). Динозавры: наиболее полная и современная энциклопедия для любителей динозавров всех возрастов. Случайный дом. Список родов для Holtz 2012 Информация о весе
- ^ Kurumada, Y .; Aoki, S .; Aoki, K .; Kato, D .; Saneyoshi, M .; Цогтбаатар, К .; Windley, B.F .; Исигаки, С. (2020). «Кальцитовый U – Pb возраст меловой формации Байн Шире, несущей позвоночных, в пустыне Восточная Гоби в Монголии: полезность каличе для определения возраста». Терра Нова. 32 (4): 246–252. Дои:10.1111 / тер.12456.
- ^ Пол, Г.С. (1984). «Сегнозавры динозавры: остатки прозауроподно-орнитисхиевого перехода?». Журнал палеонтологии позвоночных. 4 (4): 507–515. Дои:10.1080/02724634.1984.10012026. ISSN 0272-4634. JSTOR 4523011.
- ^ Занно, Л. Э .; Цогтбаатар, К .; Чинзориг, Т .; Гейтс, Т.А. (2016). «Специализации анатомии нижней челюсти и зубных рядов Segnosaurus galbinensis (Theropoda: Therizinosauria)». PeerJ. 4: e1885. Дои:10.7717 / peerj.1885. ЧВК 4824891. PMID 27069815.
- ^ Macaluso, L .; Tschopp, E .; Мэннион, П. (2018). «Эволюционные изменения ориентации лобка у динозавров сильнее коррелируют с системой вентиляции, чем с травоядными». Палеонтология. 61 (5): 703–719. Дои:10.1111 / pala.12362.
- ^ Ли, Ю. М .; Lee, H.J .; Кобаяши, Ю .; Carabajal, A. P .; Barsbold, R .; Fiorillo, A. R .; Цогтбаатар, К. (2019). «Необычное движение, сохранившееся в пределах крокодиловидной тропы из верхнемеловой формации Баяншири в Монголии, и ее палеобиологические последствия». Палеогеография, палеоклиматология, палеоэкология. 533 (109353): 2. Bibcode:2019ППП ... 533j9239L. Дои:10.1016 / j.palaeo.2019.109239.
- ^ Кубота, К .; Барсболд Р. (2007). «Новые дромеозавриды (Dinosauria Theropoda) из верхнемеловой формации Баяншири Монголии». Журнал палеонтологии позвоночных. 27 (доп. к 3): 102A. Дои:10.1080/02724634.2007.10010458.
- ^ Тернер, А. Х .; Makovicky, P.J .; Норелл, М.А. (2012). "Обзор систематики дромеозавров и паравианской филогении". Бюллетень Американского музея естественной истории. 371 (371): 1–206. Дои:10.1206/748.1. HDL:2246/6352. S2CID 83572446.
- ^ Khand, Y .; Бадамгарав, Д .; Ariunchimeg, Y .; Барсболд Р. (2000). «Меловая система Монголии и ее среды осадконакопления». Меловые среды Азии. Развитие палеонтологии и стратиграфии. 17. С. 49–79. Дои:10.1016 / s0920-5446 (00) 80024-2. ISBN 9780444502766.
- ^ Ксепка, Д. Т .; Норелл, М.А. (2006). «Эркету эллисони, длинношеий зауропод из Бор Гуве (Дорноговский аймак, Монголия)» (PDF). Американский музей Novitates. 3508 (3508): 1–16. Дои:10.1206 / 0003-0082 (2006) 3508 [1: EEALSF] 2.0.CO; 2.
















