Овираптор - Oviraptor - Wikipedia
| Овираптор | |
|---|---|
 | |
| Образец голотипа Овираптор | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Надсемейство: | †Caenagnathoidea |
| Семья: | †Oviraptoridae |
| Род: | †Овираптор Осборн, 1924 |
| Типовой вид | |
| †Овираптор филоцератопс Осборн, 1924 г. | |
| Синонимы | |
| |
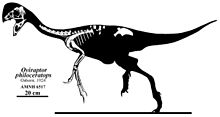
Овираптор (что означает "захватчик яиц" или "похититель яиц") род из овирапторид динозавр который жил в Азия вовремя Поздний мел период. Первые останки были собраны из Джадохтинская свита из Монголия в 1923 г. во время палеонтологической экспедиции под руководством Рой Чепмен Эндрюс, а в следующем году род и типовой вид Овираптор филоцератопс были названы Генри Фэрфилд Осборн. Название рода относится к первоначальной мысли о привычках кражи яиц, а конкретное название было предназначено, чтобы усилить эту точку зрения, указывая на предпочтение перед цератопсий яйца. Несмотря на то, что к роду отнесены многочисленные экземпляры, Овираптор известен только из одного частичного скелета, рассматриваемого как голотип, также как и гнездо около пятнадцати яиц и несколько мелких фрагментов молоди.
Овираптор был довольно маленьким пернатый oviraptorid, его длина оценивается в 1,6 м (5,2 фута), а вес составляет 33–40 кг (73–88 фунтов). У него была широкая нижняя челюсть с черепом, который, вероятно, имел гребень. И верхняя, и нижняя челюсти были беззубый и развил возбужденный клюв, который использовался при кормлении по крепкой морфологии нижней челюсти. Руки были хорошо развиты и удлинены, оканчивались тремя пальцами с загнутыми когтями. Как и другие овирапториды, Овираптор имел длинные задние конечности, четырехпалые лапы, с первым пальцем уменьшенный. Хвост, вероятно, был не очень удлинен и заканчивался пигостиль которые поддерживали большие перья.
Первоначальные отношения Овираптор были плохо изучены в то время и были отнесены к несвязанным Орнитомимиды автор оригинального описания, Генри Осборн. Однако повторные исследования, проведенные Ринчен Барсболд доказал, что Овираптор был достаточно отчетливым, чтобы оправдать отдельную семью, Oviraptoridae. При первом описании Овираптор был интерпретирован как динозавр, воровавший яйца и поедающий яйца, учитывая тесную связь голотипа с гнездом динозавров. Однако находки многочисленных овирапторозавров в позах гнездования показали, что этот экземпляр на самом деле был задумчивый гнездо и не воровать и не питаться яйцами. Более того, сообщалось об обнаружении останков небольшого птенца или птенца вместе с экземпляром голотипа, что дополнительно подтверждает родительская забота.
История открытия

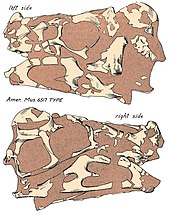
Первые остатки Овираптор были обнаружены на красноватых песчаники из Поздний мел Джадохтинская свита из Монголия, в местности Байн Дзак (также известной как Пылающие скалы ), во время палеонтологической экспедиции 1923 г. Эту экспедицию возглавил североамериканский натуралист Рой Чепмен Эндрюс и закончился открытием трех новых для науки останков теропод, в том числе останков Овираптор. Они были официально описаны североамериканским палеонтолог Генри Фэрфилд Осборн в 1924 г., который на основе нового материала назвал роды Овираптор, Saurornithoides и Велоцираптор. Особый род Овираптор возведен с типовым видом О. филоцератопс на основе голотипа AMNH 6517, частичный человек без задней части скелета, но в том числе сильно раздавленный череп, частичное шейный и спинные позвонки, грудные элементы, включая фуркула с левой рукой и частичными руками, левой подвздошная кость и немного ребра. Соответственно, этот экземпляр был найден лежащим над гнездо примерно 15 яйца - гнездо, внесенное в каталог как AMNH 6508 - с черепом, отделенным от яиц всего на 10 см (100 мм) осадок. Учитывая близость обоих образцов, Осборн интерпретировал Овираптор как динозавр с привычкой есть яйца, и объяснил, что родовое имя, Овираптор, является латинский для "захватчика яиц" или "похитителя яиц" из-за ассоциации окаменелостей. Конкретное имя, филоцератопс, означает «любовь к яйцам цератопсов», которая также возникает в результате первоначальной мысли о гнезде, относящемся к Протоцератопс или другой цератопсий. Однако Осборн предположил, что имя Овираптор могло отражать неправильное восприятие этого динозавра. Кроме того, Осборн обнаружил Овираптор быть похожими на несвязанных - в то время, однако, считавшихся связанными - быстроходных орнитомимидов на основе беззубых челюстей и назначенных Овираптор к Орнитомимиды.[1] Осборн ранее сообщал о таксоне как «Фенестрозавр филосератопс»,[2] но позже это было дискредитировано.[1]

В 1976 г. монгольский палеонтолог. Ринчен Барсболд отметил некоторые несоответствия относительно таксономического размещения Овираптор и пришел к выводу, что этот таксон совершенно отличался от орнитомимид по анатомическим признакам. Исходя из этого, он возвел Oviraptoridae содержать Овираптор и близкие родственники.[3] После первоначального описания Осборном Овирапторяйцо, связанное с голотипом, было признано принадлежащим Протоцератопс,[4][5] и овирапториды в основном считались тероподами, питающимися яйцами.[6] Тем не менее, в 1990-х годах открытие многочисленных гнездящихся и гнездящихся экземпляров овирапторид доказало, что Осборн был прав в своем предостережении относительно названия Овираптор. Эти находки показали, что овирапториды собираются и защищают свои гнезда, приседая на них. Эта новая линия доказательств показала, что гнездо связано с голотипом Овираптор принадлежал ему, и особь на самом деле вынашивала яйца в момент смерти, а не охотилась на них.[7][8][9]
Упомянутые образцы

После присвоения Oviraptoridae названия в 1976 г. Барсболд передал еще шесть экземпляров Овираптор, в том числе два частных образца под номерами MPC-D 100/20 и 100/21.[3] В 1986 году Барсболд понял, что два последних не принадлежат к роду, а представляют нового овирапторида: Конхораптор.[10] Большинство других экземпляров также вряд ли принадлежат к Овираптор сам, и они были отнесены к другим овирапторидам.[11][12] Частичная особь также с яйцами из Формация Баян Мандау Монголии в 1996 г. Дун Чжимин и Филип Дж. Карри, образец ИВПП В9608.[9] Однако в 2010 г. Николас Р. Лонгрич и два последних палеонтолога выразили свою неуверенность в отношении этого направления, поскольку существует несколько анатомических различий, таких как пропорции фаланги руки. Они пришли к выводу, что этот экземпляр был другим и неопределенным видом, не относящимся к этому таксону.[13] В 1981 году Барсболд передал образец MPC-D 100/42 в компанию Овираптор, очень хорошо сохранившаяся и довольно полная особь из джадохтинской свиты.[14] Поскольку известные элементы Овираптор были настолько фрагментарными по сравнению с другими членами, MPC-D 100/42 стал основным справочным материалом / изображением этого таксона, отмеченного как Овираптор филоцератопс в научная литература.[15][11]
Эта концепция была опровергнута Джеймс М. Кларк и его коллеги в 2002 году, которые отметили, что этот высокохохлый экземпляр имеет больше черт черепа, чем Читипати чем это происходит с Овираптор- который фактически не сохраняет гребня - и может представлять второй вид прежнего или целый новый род.[16] В 1986 году Барсболд описал второй вид Овираптор, "O. mongoliensis"на основе образца MPC-D 100 / 32a, родом из Формация Немегт.[10] Однако повторное исследование, проведенное Барсболдом в 1997 г., обнаружило достаточно различий в этом экземпляре, чтобы назвать новый род. Ринчения, но он не описал это формально, и этот новый овирапторид остался номен дубиум.[17] Это было исправлено Польский палеонтолог Halszka Osmólska и команда в 2004 г., официально назвав таксон Rinchenia mongoliensis.[18] В североамериканский палеонтолог Марк А. Норелл и коллеги в 2018 году сообщили о новом образце Овираптор: AMNH 33092, который состоит из большеберцовой кости и двух плюсневых костей птенца или очень маленького детеныша. AMNH 33092 был обнаружен вместе с голотипом и, вероятно, был частью гнезда. Овираптор теперь известен по голотипу с ассоциированными яйцами и молодью / птенцом.[19]
Описание
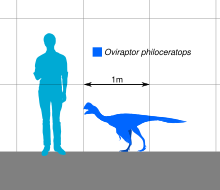
Экземпляр голотипа был оценен в длину 1,6 м (5,2 фута) при весе от 33 до 40 кг (73-88 фунтов).[20][21][22] Хотя у голотипа в основном отсутствует задняя часть скелета, вполне вероятно, что Овираптор имел две хорошо развитые задние конечности, которые заканчивались тремя функциональными пальцы ног с первым рудиментарный, а также относительно уменьшенная хвост. Как показано в связанных овирапториды руки прикрывались удлиненными перья, а хвост заканчивался пигостиль, который, как известно, поддерживает поклонника перьев.[23][24]
В череп из Овираптор был глубоким и укороченным с большим фенестры (отверстия) по сравнению с другими динозаврами, и имеет длину около 17,9 см (179 мм) в сохраненном виде. Однако фактическая длина может быть больше, учитывая, что у голотипа черепа отсутствуют некоторые области, такие как предчелюстная кость. На голотипе черепа почти полностью отсутствует гребень, однако верхние поверхности сросшихся теменных и лобных костей указывают на то, что у него, вероятно, был хорошо развитый гребень, поддерживаемый носовой и предчелюстные кости (в основном последний) из трибуна. Овираптор имели удлиненную верхнюю челюсть и зубную кость, что могло привести к более вытянутой морде по сравнению с сильно коренастыми челюстями других овирапторид. В нёбо жесткий, продолжается ниже линии челюсти и образован предчелюстными костями, сошники, и максиллы. Как и у других овирапторид, он мог иметь пару зубчатых выступов на небе, направленных вниз. Как и у других овирапторид, ноздри (внешние ноздри) были бы относительно небольшими и располагались высоко на черепе. Овираптор имел беззубый челюсти, которые заканчивались крепким, попугай -подобно рамфотека (роговой клюв). Кривизна кончика зубной кости была опущена, но менее выражена, чем у других овирапторид, таких как Читипати. В целом нижняя челюсть представляет собой короткую и глубокую кость длиной 19,5 см (195 мм).[16]

Как и у большинства овираптордов, нервные отростки голотипа шейные позвонки позвонки были короткими, а нервные дуги были Х-образными. Однако шипы на задних позвонках становятся более выраженными. Зигапофизы первых шейных позвонков расположены параллельно друг другу, а постзигапофизы, по-видимому, существенно не расходятся от средней линии, в основном аналогично Читипати. В шейные ребра на голотипе срослись с позвонками.[25][19] Нервные отростки в переднем ряду позвоночника имеют прямоугольную форму. спинные позвонки при осмотре сбоку и больше, чем шипы шейных позвонков. На переднем переднем спинном позвонке можно найти несколько плевроцелей (небольших отверстий с воздушным зазором), похожих на плевроцели. Khaan.[25]
В фуркула из Овираптор очень отличается от других овирапторид тем, что имеет киль по средней линии на передней поверхности гипоклеидия - направленный вниз выступ в центре фуркулы. Кость V-образная, в поперечном сечении округлая, с удлиненным шиповидным гипоклеидием, межключичный угол около 90 °.[26] В скапулокоракоид слита в голотипе, однако коракоидный сильно поврежден. В лопатка слегка изогнут и имеет длину 23 см (230 мм). Овираптор имел относительно удлиненную руку, состоящую из плечевая кость, радиус, локтевая кость, и манус. Фаланговая формула Овираптор было 2-3-4, как и у большинства других теропод и овирапторид. Рука Овираптор имел три тонких и похожих на птицу пальца, каждый из которых был сплющен и загнут из стороны в сторону ногтевые (коготь кость). В отличие от некоторых овирапторид, Овираптор не пострадал уменьшения второго и третьего пальца относительно первого.[1][25][27]
Считается несовершеннолетним Овираптор AMNH 33092 сохраняет материал задних конечностей, включающий правую большеберцовую кость с III и IV плюсневыми костью. Его большеберцовая кость имеет длину 5,87 см (58,7 мм), что свидетельствует о значительно меньших размерах, чем голотип.[19] Гнездо AMNH 6508 консервы элонгатоолитид яйца, каждое из которых имеет длину 14 см (140 мм) (некоторые из них неполные). Тем не менее, существует вероятность того, что тафономическое дробление могло сжать их до 2 см (20 мм).[7][28]
Классификация
Овираптор изначально был в союзе с орнитомимиды Осборна из-за его беззубого клюва. Осборн также обнаружил сходство с Хиростеноты, который до сих пор считается близким родственником Овираптор.[1] В 1976 г. Барсболд построил новую семью, чтобы содержать Овираптор и его близкие родственники, делая Овираптор то типовой род из Oviraptoridae.[3] Во время переописания голотипа черепа Кларком и его коллегами в 2002 году они отметили, что Овираптор имели относительно удлиненные верхнечелюстную и зубную кость. Эти черты менее выражены у производных овирапторид и предполагают, что Овираптор принадлежит к ближнему основанию Oviraptoridae.[16]
В кладограмма ниже следует анализ Грегори Ф. Фанстон и коллеги в 2020 году:[29]
| Oviraptoridae |
| ||||||||||||||||||||||||
Палеобиология
Кормление

Впервые описанный Осборном в 1924 году, Овираптор изначально предполагалось, что яйцекладущий - организм, который придерживается яичной диеты - образ жизни, основанный на ассоциации голотипа с гнездом, которое, как считается, принадлежит Протоцератопс.[1] В 1977 году Барсболд выдвинул гипотезу сокрушительной челюсти. Он утверждал, что сила крепкой нижней челюсти и, вероятно, рамфотека (рогатый клюв) был достаточно сильным, чтобы сломать панцири моллюски Такие как моллюски, которые находятся в той же геологической формации, что и Овираптор. Эти кости являются частью основной кости верхней челюсти или верхняя челюсть, которые сходятся посередине, образуя пару зубцов. Рамфотека и нижняя челюсть вместе с продолжением нескольких костей от нёбо, сделал бы пирсинг. Барсболд также предположил, что овирапториды могли иметь полуводный образ жизни, основанный на диете на основе моллюсков, высоком расположении носовых полостей, увеличенной мускулатуре хвоста и большем размере первого руководства цифра.[30] В тезисе конференции 1990 года Дэвид К. Смит представил остеологическую переоценку Овираптор где он отверг заявления Барсболда. Он не нашел никаких доказательств того, что передняя конечность специализировалась на водное движение, и челюсти, вместо того чтобы сохранять механизм дробления, сохраняют поверхности среза. Как череп беззубый, легкого телосложения и не имеющего нескольких сильных областей прикрепления мышц, Смит предположил, что листья могли быть важной частью диеты Овираптор.[31] Однако в 1995 году Норелл и его коллеги сообщили о фрагментированных останках ящерица в полости тела экземпляра голотипа, предполагая, что Овираптор был частично плотоядный.[32]

В 2008 году Стиг Олав К. Янсен сравнил черепа нескольких видов овирапторид с черепами птицы и черепахи исследовать, какие свойства могут предсказать наличие рамфотеки. Он обнаружил, что нижние челюсти овирапторид очень похожи на челюсти. попугаи, а верхняя челюсть больше похожа на черепаху. Основываясь на этих наблюдениях, Янсен предположил, что овирапториды были всеядный поскольку резко развитая рамфотека вместе с выступающими передними конечностями была бы приспособлена для захвата и отрыва небольших добыча. Более того, заостренные выступы неба помогли удерживать добычу. Янсен отметил, что полностью травоядный диета овирапторид кажется маловероятной, поскольку им не хватало плоских и широких Томия (режущие края нижних челюстей) до жевать, и не могли двигать нижними челюстями в сторону. Однако он считал нижнюю челюсть достаточно сильной, чтобы в ней могли быть по крайней мере раздавленные элементы, такие как яйца, орехи или другой жесткий семена.[33]
Лонгрич и его коллеги в 2010 году также отвергли дурофаг (животные, практикующие дробление панцирей) гипотеза, учитывая, что такие животные обычно развивают зубы с широкими поверхностями измельчения. Заостренная форма зубных костей нижней челюсти позволяет предположить, что у овирапторид вместо этого для стрижки пищи использовалась остроконечная рамфотека. В симфизарный Область (соединение кости) в передней части зубной кости могла дать некоторую способность к раздавливанию, но, поскольку это была относительно небольшая область, вероятно, это не была основная функция челюстей. Еще одним аргументом против того, что они поедали моллюсков, является тот факт, что большинство овирапторид были обнаружены в отложения которые интерпретируются как представляющие преимущественно засушливые или полузасушливые среды, такие как Овираптор в Джадохтинская свита. Команда также обнаружила, что овирапториды и дицинодонты общие черты черепа, такие как короткие, глубокие и беззубые нижние челюсти; удлиненные зубные симфизы; удлиненные нижнечелюстные отверстия; и острое небо. Современные животные с челюстями, напоминающими челюсти овирапторид, включают попугаев и черепахи; последняя группа также имеет зубчатые выступы на небной области. Лонгрич и его коллеги пришли к выводу, что из-за сходства между овирапторидами и травоядными животными основная часть их рациона, скорее всего, была сформирована из растительного материала. Челюсти овирапторид могли быть специализированы для обработки пищи, например ксерофитный растительность - растительность, приспособленная к окружающей среде с небольшим количеством воды - которая могла бы расти в их засушливой среде, но это невозможно продемонстрировать, так как мало что известно о палеофлоре пустыня Гоби.[13]
Однако в 2018 году Фунстон и его коллеги поддержали гипотезу сокрушительной челюсти. Они отметили, что коренастый трибуна а крепкие нижние челюсти овирапторид предполагают, на самом деле, сильный и кусающий укус, который очень похож на укус попугаев. Фунстон и его коллеги считали эти анатомические особенности овирапторид совместимыми с плодоядный диета, включающая орехи и семена.[34]
Размножение


Поскольку описание эмбриональный Читипати Образец в 1994 году овирапторид стал более понятым: вместо того, чтобы питаться яйцами, они на самом деле размышляли и заботились о гнезда. Этот образец показал, что голотип Овираптор вероятно был половозрелый человек, который погиб инкубация связанное гнездо с яйца.[7] Это новое поведение овирапторид стало более ясным после отчета и краткого описания взрослой особи гнездящейся особи. Читипати в 1995 году Норелл и его коллеги. Образец был найден на вершине яичные кладки, при этом его задние конечности симметрично согнуты по обе стороны от гнезда, а передние конечности покрывают периметр гнезда. Эта задумчивая поза встречается сегодня только у современные птичьи динозавры и поддерживает поведенческую связь между последней группой и нептичьи динозавры.[8] В 1996 году Донг и Карри описали новый экземпляр гнездящихся овирапторид из Формация Баян Мандау. Он был найден лежащим на вершине гнезда, состоящего примерно из 6 яиц, которые сохранились, и они были заложены в форме холма с круговым рисунком. Поскольку образец был найден над гнездом, его передние конечности закрывали яйца, а частично сохранившаяся задняя лапа - около центра гнезда, Донг и Карри предположили, что он был пойман и захоронен песчаная буря во время инкубации. Они исключили возможность того, что овирапториды могут похитить яйца, поскольку они либо съели, либо инстинктивно покинули гнездо задолго до того, как оно было похоронено песчаной бурей или другой. метеорологическое явление.[9]
В 1999 году Кларк и его команда подробно описали ранее сообщенное Читипати гнездящегося экземпляра и кратко обсудили образец голотипа Овираптор и его связь с гнездом AMNH 6508. Они отметили, что точное положение, в котором голотип был найден над гнездом, неясно, поскольку они были разделены во время подготовка Гнездо не полностью завершено, сохранилось около 15 яиц, два из которых повреждены. Более того, полукруглое расположение гнезда указывает на то, что яйца откладывались парами и по крайней мере в три кольца, и это гнездо изначально было круглым, подобно гнезду. курган.[28]
Томас П. Хопп и Марк Дж. Орсен в 2004 году проанализировали насиживающее поведение вымерших и существующих видов динозавров, включая овирапторид, чтобы оценить причину удлинения и развития крыльев и хвоста. перья. Учитывая, что самый полный экземпляр гнездящихся овирапторид - в то время, 1995 г. Читипати гнездящийся экземпляр - был обнаружен в позе, очень похожей на птичью, с передними конечностями в почти сложенном положении, а грудная часть, живот и ступни соприкасались с яйцами, Хопп и Орсен указали, что Pennaceous перья и перьевое покрытие скорее всего присутствовали в жизни. «Крылья» и хвост овирапторид обеспечили бы защиту яиц и детеныши против климатических факторов, таких как Солнечный свет, ветер, и идет дождь. Однако руки этого экземпляра не были сильно сложены, как у некоторых современных птиц, а более вытянуты, напоминая стиль большого размера. нелетающие птицы словно страус. Вытянутая рука также похожа на поведение этой птицы, которая, как известно, гнездится большими кладками, такими как овирапториды. Основываясь на положении передних конечностей гнездящихся овирапторид, Хопп и Орсен предложили насиживание как наследственную причину удлинения крыльев и хвостовых перьев, поскольку была большая потребность в обеспечении оптимального защита для яиц и молоди.[35]

В 2005 году Тамаки Сато и его команда сообщили о необычном образце овирапторида из Формация Наньсюн. Этот новый образец, как было обнаружено, сохранил в основном тазовую область с двумя яйцами внутри, что указывает на женский. Размер и положение яиц позволяют предположить, что овирапториды сохранили два функциональных яйцеводы, но уменьшил количество яиц овулированный по одному на яйцевод.[36] Дэвид Дж. Варриккио и его коллеги в 2008 году обнаружили, что относительно большой размер яйцекладки овирапторид и троодонтиды больше всего похож на птиц современных птиц, которые практикуют полигамный вязка и обширный самец родительская забота, Такие как ратит птицы, предполагающие схожие повадки. Этот репродуктивная система скорее всего, представляет собой наследственное состояние современных птиц, а забота о двух родителях (в которой участвуют оба родителя) является более поздним развитием.[37] В 2014 г. В. Скотт Персонс и его коллеги предположили, что овирапторозавры вторично нелетают, и некоторые черты их хвостов могут указывать на склонность к поведение дисплея, Такие как дисплей ухаживания. Хвост нескольких овирапторозавров и овирапторид заканчивался пигостили, костная структура на конце хвоста, которая, по крайней мере, у современных птиц, используется для поддержки веера из перьев. Кроме того, хвост был особенно мускулистым и обладал ярко выраженной гибкостью, что, возможно, помогало при ухаживании.[24]

В 2018 году Цзы-Руэй Ян и его коллеги определили кутикула слои на нескольких яичных скорлупах динозавров манирапторанов, в том числе и овирапторид. Эти конкретные слои состоят из белки, полисахариды и пигменты, но в основном из липиды и гидроксиапатит. У современных птиц они служат для защиты яиц от обезвоживание и вторжение микроорганизмы. Поскольку большинство экземпляров овирапторид были обнаружены в образованиях Caliche Янг и его коллеги предположили, что яйца, покрытые кутикулой, были репродуктивной стратегией, адаптированной для повышения их успешности вылупления в таком засушливом климате и окружающей среде.[38]
В 2019 году Ян и его коллеги пересмотрели гипотезу терморегулирующий контактная инкубация с использованием полных гнезд овирапторид из формации Nanxiong и предоставила детальную реконструкцию архитектуры кладки овирапторид. Они отметили, что взрослые особи овирапторид, связанные с гнездом, не обязательно инкубировали яйца, поскольку они могли представлять самку в процессе откладки яиц, а многокольцевое сцепление препятствовало достаточной теплопередача от родителя к внутренним кольцам яиц. Обычное гнездо овирапторид было построено в виде полого наклонного холма с высокоорганизованной архитектурой: яйца, вероятно, были пигментированы и располагались парами, каждая пара располагалась в виде трех-четырех эллиптических колец. Поскольку родитель, вероятно, действовал из центра гнезда, в этой области не было яиц. Ян и его коллеги пришли к выводу, что стиль гнездования овирапторид настолько уникален, что им не хватает современных аналогов, поэтому использование репродукции овирапторид может быть не лучшим примером для информирования об эволюции репродуктивных стратегий птиц. Однако команде не удалось определить, был ли несовершеннолетний Овираптор AMNH 33092 вылупился из гнезда, связанного с голотипом.[39]
Палеосреда
Овираптор известен из урочища Байн-Дзак на Джадохтинская свита в Монголии образование, восходящее к Поздний мел примерно от 71 миллиона до 75 миллион лет назад.[40] Палеосреда джадохтинской свиты интерпретируется как имеющая полузасушливый климат, с песчаная дюна и аллювиальный настройки похожи на современные пустыня Гоби. Полузасушливый степь пейзаж был осушен прерывистые потоки и иногда подвергался воздействию пыли и песчаные бури, влажность была сезонной.[41][42] Хотя считается, что это образование в значительной степени относится к сохранившимся сильно засушливым условиям, некоторые недолговечные водоемы поступили из местонахождения Ухаа Толгод, по данным речной седиментация.[43] Кроме того, считается, что позже в Кампанский возраст и в Маастрихтский, климат сместится к более влажной речной среде, наблюдаемой в Формация Немегт.[42]
Джадохтинская свита разделяется на нижнюю байн-дзакскую пачку и верхнюю тургругинскую пачку. Известные останки Овираптор были произведены членом Байн Дзак, который также дал динозавров Байноцератопс, Пинакозавр, Протоцератопс, Saurornithoides, Велоцираптор,[40] и Halszkaraptor.[44] Дальнейшая фауна динозавров из этой пачки включает фауну местонахождения Ухаа Толгод, состоящую из Апсаравис, Байронозавр, Читипати, Гобиптерикс, Khaan, Хол, Шуувуя, Цааган,[43] и Минотауразавр.[45]
Тафономия
Поза голотипа Овираптор наряду с ассоциацией яиц, предполагают, что они застряли над гнездом во время песчаная буря, и захоронение был относительно быстрым, учитывая, что у тела не было возможности полностью разъединиться или очищенный хищниками.[1][9] Палеонтолог Кеннет Карпентер также согласились с тем, что песчаные бури, возможно, были наиболее вероятным случаем захоронения яиц, найденных в отложениях.[46] Среди элементов череп стал особенно уплощенным и искаженным во время процесс окаменения.[16]
Смотрите также
Рекомендации
- ^ а б c d е ж Осборн, Х. Ф. (1924). «Три новых теропода, зона Protoceratops, центральная Монголия». Американский музей Novitates (144): 1−12. HDL:2246/3223. OCLC 40272928.
- ^ Осборн, Х. Ф. (1924). «Открытие неизведанного континента». Естественная история. 24 (2): 133−149.
- ^ а б c Барсболд Р. (1976). "О новом позднемеловом семей- стве мелких теропод Oviraptoridae fam. Nov Монголии" [Новое позднемеловое семейство мелких тероподы Oviraptoridae n. fam. в Монголии]. Доклады Академии Наук СССР. 226 (3): 685−688.
- ^ Кумбс, У. П. (1989). «Современные аналоги гнездования и родительского поведения динозавров». В Фарлоу, Дж. О. (ред.). Палеобиология динозавров. Специальный доклад Геологического общества Америки 238. Колорадо: Боулдер. С. 21−54. Дои:10.1130 / SPE238-p21.
- ^ Талборн, Р. А. (1992). «Гнездо динозавра Протоцератопса». Lethaia. 25 (2): 145−149. Дои:10.1111 / j.1502-3931.1992.tb01379.x.
- ^ Карри, П. Дж .; Годфри, С. Дж .; Несов, Л. А. (1993). «Новые образцы ценагнатидов (Dinosauria: Theropoda) из верхнего мела Северной Америки и Азии». Канадский журнал наук о Земле. 30 (10): 2255−2272. Bibcode:1993CaJES..30.2255C. Дои:10.1139 / e93-196.
- ^ а б c Норелл, М. А .; Clark, J.M .; Дашзевег, Д .; Barsbold, R .; Chiappe, L.M .; Дэвидсон, А. Р .; McKenna, M.C .; Altangerel, P .; Новачек, М. Дж. (1994). "Эмбрион динозавра-теропод и сходство яиц динозавров из Пламенных Скал". Наука. 266 (5186): 779−782. Bibcode:1994Наука ... 266..779N. Дои:10.1126 / science.266.5186.779. JSTOR 2885545. PMID 17730398.
- ^ а б Норелл, М. А .; Clark, J.M .; Chiappe, L.M .; Дашзевег, Д. (1995). «Гнездовой динозавр». Природа. 378 (6559): 774−776. Bibcode:1995Натура 378..774Н. Дои:10.1038 / 378774a0.
- ^ а б c d Dong, Z .; Карри, П. Дж. (1996). «Об обнаружении скелета овирапторид в гнезде из яиц в Баян Мандаху, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле. 33 (4): 631−636. Bibcode:1996CaJES..33..631D. Дои:10.1139 / e96-046.
- ^ а б Барсболд Р. (1986). "Хищные динозавры овирапторы". В Воробьевой, Е. И. (ред.). Герпетологические исследования в Монгольской Народной Республике (на русском). Институт эволюционной морфологии и экологии животных: Доклады Академии Наук СССР. С. 210–223.
- ^ а б Марьянская, Т .; Osmólska, H .; Волсан, М. (2002). «Статус Авиалана для Овирапторозаврии» (PDF). Acta Palaeontologica Polonica. 47 (1): 97−116.
- ^ Kundrát, M .; Яначек, Ю. (2007). «Пневматизация черепа и слуховое восприятие овирапторидного динозавра Conchoraptor gracilis (Theropoda, Maniraptora) из позднего мела Монголии». Naturwissenschaften. 94 (9): 769−778. Bibcode:2007NW ..... 94..769R. Дои:10.1007 / s00114-007-0258-7.
- ^ а б Longrich, N.R .; Карри, П. Дж .; Донг, З. (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандаху, Внутренняя Монголия». Палеонтология. 53 (5): 945−960. Дои:10.1111 / j.1475-4983.2010.00968.x.
- ^ Барсболд Р. (1981). "Беззубые хищные динозавры Монголии" [Беззубые хищные динозавры Монголии]. Труды совместной советско-монгольской палеонтологической экспедиции (на русском). 15: 28−39. S2CID 195060070. Переведенная статья
- ^ Barsbold, R .; Марьянская, Т .; Осмольска, Х. (1990). «Овирапторозаврия». В Weishampel, D. B .; Osmolska, H .; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли: Калифорнийский университет Press. С. 249−258. ISBN 9780520067271.
- ^ а б c d Clark, J.M .; Норелл, М. А .; Роу, Т. (2002). "Анатомия черепа Citipati osmolskae (Theropoda, Oviraptorosauria) и переосмысление голотипа Oviraptor philoceratops" (PDF). Американский музей Novitates (3364): 1−24. Дои:10.1206 / 0003-0082 (2002) 364 <0001: CAOCOT> 2.0.CO; 2. HDL:2246/2853.
- ^ Барсболд, 1997. Овирапторозаврия. У Карри и Падиана (ред.). Энциклопедия динозавров. 505-509.
- ^ Osmólska, H .; Карри, П. Дж .; Барсболд Р. (2004). «Овирапторозаврия». В Weishampel, D. B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 165−183. ISBN 0520242092.
- ^ а б c Норелл, М. А .; Баланов, А. М .; Barta, D.E .; Эриксон, Г. М. (2018). «Второй экземпляр Citipati osmolskae, связанный с гнездом яиц из Ухаа Толгод, Омногов Аймаг, Монголия». Американский музей Novitates (3899): 1−44. HDL:2246/6858.
- ^ Werner, J .; Грибелер, Э. М. (2013). «Новые взгляды на воспроизводство нептичьих динозавров и их эволюционные и экологические последствия: связь ископаемых свидетельств с аллометрией существующих близких родственников». PLOS ONE. 8 (8): e72862. Дои:10.1371 / journal.pone.0072862. ЧВК 3749170. PMID 23991160.
- ^ Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 178. ISBN 9780691167664.
- ^ Кампионе, Н.Е .; Эванс, Д. К. (2020). «Точность и точность оценки массы тела у нептичьих динозавров». Биологические обзоры. Дои:10.1111 / brv.12638.
- ^ Fanti, F .; Карри, П. Дж .; Бадамгарав, Д .; Лалуэза-Фокс, К. (2012). «Новые образцы немегтомайи из свит Баруунгойот и Немегт (поздний мел) Монголии». PLOS ONE. 7 (2): e31330. Bibcode:2012PLoSO ... 731330F. Дои:10.1371 / journal.pone.0031330. ЧВК 3275628. PMID 22347465.
- ^ а б Лица, W. S .; Карри, П. Дж .; Норелл, М.А. (2014). «Формы и функции хвоста овирапторозавра». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0093.
- ^ а б c Баланов, А. М .; Норелл, М.А. (2012). «Остеология Khaan mckennai (Oviraptorosauria, Theropoda)». Бюллетень Американского музея естественной истории (372): 1−77. Дои:10.1206/803.1. HDL:2246/6413. OCLC 884738416.
- ^ Nesbitt, S.J .; Тернер, А. Х .; Сполдинг, М .; Conrad, J. L .; Норелл, М.А. (2009). "Теропод фуркула". Журнал морфологии. 270 (7): 856−879. Дои:10.1002 / jmor.10724. PMID 19206153.
- ^ Lü, J .; Карри, П. Дж .; Xu, L .; Чжан, X .; Pu, H .; Цзя, С. (2013). «Овирапторидные динозавры размером с курицу из центрального Китая и их онтогенетические последствия» (PDF). Naturwissenschaften. 100 (2): 165−175. Bibcode:2013NW .... 100..165л. Дои:10.1007 / s00114-012-1007-0. PMID 23314810.
- ^ а б Clark, J.M .; Норелл, М. А .; Чиаппе, Л. М. (1999). «Скелет овирапторид из позднего мела Ухаа Толгод, Монголия, сохранившийся в птичьем положении для размышлений над гнездом овирапторид». Американский музей Novitates (3265): 1−36. HDL:2246/3102.
- ^ Funston, G.F .; Цогтбаатар, Ц .; Цогтбаатар, К .; Кобаяши, Ю .; Салливан, К .; Карри, П. Дж. (2020). «Новый двупалый динозавр проливает свет на излучение Овирапторозаврии». Королевское общество открытой науки. 7 (10): 201184. Дои:10.1098 / rsos.201184.
- ^ Барсболд Р. (1977). «Кинетизм и особенности челюстного аппарата овирапторов (Theropoda, Saurischia)». Советско-Монгольская палеонтологическая экспедиция, Труды.. 4: 37−47. Переведенная статья
- ^ Смит, Д. К. (1990). «Остеология Oviraptor philoceratops, возможного травоядного теропод из верхнего мела Монголии». Журнал палеонтологии позвоночных. 10 (Supp. 003): 42A. Дои:10.1080/02724634.1990.10011841.
- ^ Норелл, М. А .; Gaffney, E. S .; Дингус, Л. (1995). Открытие динозавров в Американском музее естественной истории. Knopf Inc. стр. 225.
- ^ Янсен, С. О. К. (2008). Морфология клюва у овирапторид на основе современных птиц и черепах (Тезис). Университет Осло. С. 1−48. HDL:10852/11785.
- ^ Funston, G.F .; Mendonca, S.E .; Карри, П. Дж .; Barsbold, R .; Барсболд Р. (2018). «Анатомия, разнообразие и экология овирапторозавров в бассейне Немегт». Палеогеография, палеоклиматология, палеоэкология. 494: 101−120. Bibcode:2018ППП ... 494..101Ф. Дои:10.1016 / j.palaeo.2017.10.023.
- ^ Hopp, T. P .; Орсен, М. Дж. (2004). "Наследственное поведение динозавров и происхождение перьев полета" (PDF). В Currie, P. J .; Koppelhus, E.B .; Шугар, М. А .; Райт, Дж. Л. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам. Блумингтон: Издательство Индианского университета. С. 234-250.
- ^ Сато, Т .; Cheng, Y.-N .; Wu, X.-C .; Зеленицкий, Д.К .; Сяо, Ю.-Ф. (2005). "Пара яиц в скорлупе внутри самки динозавра" (PDF). Наука. 308 (5720): 375. Дои:10.1126 / наука.1110578. PMID 15831749. S2CID 19470371.
- ^ Varricchio, D. J .; Мур, Дж. Р .; Эриксон, Г. М .; Норелл, М. А .; Джексон, Ф. Д .; Борковский, Дж. Дж. (2008). "Птичий отцовский уход произошел от динозавров". Наука. 322 (5909): 1826−1828. Bibcode:2008Научный ... 322.1826V. Дои:10.1126 / science.1163245. PMID 19095938.
- ^ Yang, T.-R .; Chen, Y.-H .; Wiemann, J .; Spiering, B .; Сандер, П. М. (2018). "Ископаемая кутикула яичной скорлупы проясняет экологию гнездования динозавров". PeerJ. 6: e5144. Дои:10.7717 / peerj.5144. ЧВК 6037156. PMID 30002976.
- ^ Yang, T.-R .; Wiemann, J .; Xu, L .; Cheng, Y.-N .; Wu, X.-C .; Сандер, П. М. (2019). «Реконструкция кладок овирапторид проливает свет на их уникальную биологию гнездования». Acta Palaeontologica Polonica. 466: 581−596. Дои:10.4202 / app.00497.2018.
- ^ а б Дашзевег, Д .; Dingus, L .; Loope, D. B .; Swisher III, C.C .; Dulam, T .; Суини, М. Р. (2005). «Новое стратиграфическое подразделение, обстановка осадконакопления и оценка возраста джадохтинской свиты верхнего мела, Южный бассейн Улан-Нур, Монголия» (PDF). Американский музей Novitates (3498): 1−31. Дои:10.1206 / 0003-0082 (2005) 498 [0001: NSSDEA] 2.0.CO; 2. HDL:2246/5667.
- ^ Ежикевич, Т. (1997). «Джадохтинская свита». В Currie, P. J .; Падиан, К. (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. стр.188 −191. ISBN 978-0-12-226810-6.
- ^ а б Чинзориг, Т .; Кобаяши, Ю .; Цогтбаатар, К .; Карри, П. Дж .; Watabe, M .; Барсболд Р. (2017). "Первый орнитомимид (Theropoda, Ornithomimosauria) из верхнемеловой джадохтинской формации Тогрёгийн Шири, Монголия". Научные отчеты. 7 (5835). Bibcode:2017НатСР ... 7.5835C. Дои:10.1038 / s41598-017-05272-6. ЧВК 5517598. PMID 28724887.
- ^ а б Dingus, L .; Loope, D. B .; Дашзевег, Д .; Swisher III, C.C .; Минджин, К .; Novacek, M. J .; Норелл, М.А. (2008). «Геология Ухаа Толгод (Джадохтинская свита, верхний мел, бассейн Немегт, Монголия)» (PDF). Американский музей Novitates (3616): 1−40. Дои:10.1206/442.1. HDL:2246/5916.
- ^ Cau, A .; Бейранд, В .; Voeten, D. F. A. E .; Фернандес, В .; Tafforeau, P .; Stein, K .; Barsbold, R .; Цогтбаатар, К .; Карри, П. Дж .; Годфройт, П. (2017). «Синхротронное сканирование выявляет экоморфологию амфибий в новой стае птицеподобных динозавров». Природа. 552 (7685): 395−399. Bibcode:2017Натура.552..395C. Дои:10.1038 / природа24679. PMID 29211712.
- ^ Alicea, J .; Лёвен, М. (2013). «Новый материал Minotaurasaurus из формации Джодокта устанавливает новые таксономические и стратиграфические критерии для таксона». Журнал палеонтологии позвоночных. Программа и аннотации: 76.
- ^ Карпенер, К. (1999). «Яйца как ископаемые». Яйца, гнезда и детеныши динозавров: взгляд на воспроизводство динозавров (жизнь прошлого). Блумингтон: Издательство Индианского университета. п. 111. ISBN 9780253334978.
внешняя ссылка
 СМИ, связанные с Овираптор в Wikimedia Commons
СМИ, связанные с Овираптор в Wikimedia Commons Данные, относящиеся к Овираптор в Wikispecies
Данные, относящиеся к Овираптор в Wikispecies- Овираптор гнездо AMNH 6508 фотографий на AMNH
- Овираптор голотип черепа фотография на AMNH
















