Глубоководная рыба - Deep sea fish

Глубоководная рыба находятся рыбы живущие в темноте под освещенными солнцем поверхностными водами, то есть ниже эпипелагический или фотическая зона из море. В рыба-фонарь это, безусловно, самая распространенная глубоководная рыба. Другой глубокое море рыбы включают фонарик рыба, резак акула, щетинистый рот, удильщик, гадюка, и некоторые виды бельдюга.
Только около 2% известных морских видов обитают в пелагический Окружающая среда. Это означает, что они живут в столб воды в отличие от бентосный организмы, обитающие на морском дне или на его поверхности.[1] Глубоководные организмы обычно обитают батипелагический (Глубина 1000–4000 м) и абиссопелагический (Глубина 4000–6000 м) зоны. Однако характеристики глубоководных организмов, такие как биолюминесценция можно увидеть в мезопелагический (Глубина 200–1000 м). Мезопелагическая зона - это дифотическая зона, что означает, что свет там минимален, но все же измерим. Слой минимального содержания кислорода существует где-то на глубине от 700 до 1000 м в зависимости от места в океане. В этой области также больше всего питательных веществ. В батипелагический и абиссопелагический зоны афотический, что означает, что в эту часть океана не проникает свет. Эти зоны составляют около 75% обитаемого пространства океана.[2]
Зона эпипелагиали (0–200 м) - это область, где свет проникает в воду и происходит фотосинтез. Это также известно как фотическая зона. Поскольку обычно он простирается всего на несколько сотен метров под водой, глубокое море, около 90% объема океана, находится в темноте. Глубокое море также является чрезвычайно враждебной средой, с температурами, которые редко превышают 3 ° C (37,4 ° F) и опускаются до -1,8 ° C (28,76 ° F) (за исключением экосистем гидротермальных источников, которые могут превышать 350 ° C). C или 662 ° F), низкий уровень кислорода и давление от 20 до 1000 атмосферы (от 2 до 100 мегапаскали ).[3]
Окружающая среда

В глубоком океане воды простираются намного ниже эпипелагиали и служат местом обитания самых разных видов пелагических рыб, приспособленных к жизни в этих более глубоких зонах.[4]
В глубокой воде, морской снег это непрерывный поток органических детрит падение из верхних слоев водной толщи. Его происхождение лежит в деятельности внутри производственной фотическая зона. Морской снег включает мертвые или умирающие планктон, протисты (диатомеи ), фекалии, песок, сажа и прочая неорганическая пыль. «Снежинки» со временем растут и могут достигать нескольких сантиметров в диаметре, путешествуя неделями, прежде чем достичь дна океана. Однако большинство органических компонентов морского снега потребляются микробы, зоопланктон и другие фильтрующие животные в пределах первых 1000 метров пути, то есть в пределах эпипелагической зоны. Таким образом, морской снег можно считать основой глубоководных мезопелагический и бентосный экосистемы: Поскольку солнечный свет не может достичь их, глубоководные организмы в значительной степени полагаются на морской снег как источник энергии. Поскольку в глубоком море нет света (афотический), не хватает первичных продуцентов. Таким образом, большинство организмов в батипелагических условиях полагаются на морской снег из регионов, расположенных выше по вертикали.
Некоторые глубоководные пелагические группы, такие как рыба-фонарь, риджхед, морской топорик, и светлая рыба семьи иногда называют псевдокеанский потому что вместо того, чтобы иметь равномерное распределение в открытой воде, они встречаются в значительно большей численности вокруг структурных оазисов, особенно подводные горы и более континентальные склоны. Это явление объясняется также обилием видов жертв, которых также привлекают эти строения.
Гидростатическое давление увеличивается на 1 атмосферу на каждые 10 м глубины.[5] Глубоководные организмы испытывают такое же давление внутри своего тела, какое оказывается на них извне, поэтому они не подвержены сильному давлению. Однако их высокое внутреннее давление приводит к снижению текучести их мембран, поскольку молекулы сжимаются вместе. Текучесть клеточных мембран увеличивает эффективность биологических функций, в первую очередь выработки белков, поэтому организмы адаптировались к этому обстоятельству, увеличивая долю ненасыщенных жирных кислот в липидах клеточных мембран.[6] Помимо различий во внутреннем давлении, эти организмы выработали другой баланс между своими метаболическими реакциями, чем те организмы, которые живут в эпипелагической зоне. Дэвид Уортон, автор Жизнь на пределе возможностей: организмы в экстремальных условиях, примечания «Биохимические реакции сопровождаются изменениями объема. Если реакция приводит к увеличению объема, она будет подавлена давлением, тогда как, если она связана с уменьшением объема, она будет усилена».[7] Это означает, что их метаболические процессы должны в конечном итоге до некоторой степени уменьшать объем организма.
Большинство рыб, которые эволюционировали в этой суровой окружающей среде, не способны выжить в лабораторных условиях, и попытки удержать их в неволе привели к их гибели. Глубоководные организмы содержат заполненные газом пространства (вакуоли).[9] Газ сжимается под высоким давлением и расширяется под низким давлением. Из-за этого известно, что эти организмы взрываются, если выходят на поверхность.[7]
Характеристики
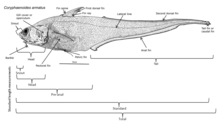



Глубоководные рыбы приобрели различные приспособления, чтобы выжить в этом регионе. Поскольку многие из этих рыб обитают в регионах, где нет естественных освещение, они не могут полагаться только на свое зрение для обнаружения добычи и партнеров и избегания хищников; глубоководная рыба развился соответственно крайнему субфотическому региону, в котором они живут. Многие из этих организмов слепы и полагаются на свои другие органы чувств, такие как чувствительность к изменениям местного давления и запаха, чтобы поймать пищу и избежать ее поимки. У тех, кто не слеп, большие и чувствительные глаза, которые могут использовать биолюминесцентный свет. Эти глаза могут быть в 100 раз более чувствительными к свету, чем человеческие глаза. Родопсин (Rh1) - это белок, содержащийся в палочковидных клетках глаза, который помогает животным видеть при тусклом свете. Хотя у большинства позвоночных обычно есть один ген опсина Rh1, некоторые глубоководные рыбы имеют несколько генов Rh1 и один вид, серебряный колючий плавник (Diretmus argenteus), имеет 38.[10] Такое распространение генов Rh1 может помочь глубоководным рыбам видеть глубины океана. Кроме того, чтобы избежать нападения хищников, многие виды имеют темный цвет, чтобы сливаться с окружающей средой.[11]
Многие глубоководные рыбы биолюминесцентный с очень большими глазами, приспособленными к темноте. Биолюминесцентные организмы способны производить свет биологическим путем за счет возбуждения молекул люциферина, которые затем производят свет. Этот процесс необходимо проводить в присутствии кислорода. Эти организмы распространены в мезопелагиали и ниже (200 м и ниже). Более 50% глубоководных рыб, а также некоторые виды креветок и кальмаров способны к биолюминесценции. Около 80% этих организмов имеют фотофоры - железистые клетки, производящие свет, которые содержат светящиеся бактерии, окаймленные темной окраской. Некоторые из этих фотофоров содержат линзы, похожие на линзы в глазах людей, которые могут усиливать или уменьшать излучение света. Способность производить свет требует только 1% энергии организма и имеет множество целей: он используется для поиска пищи и привлечения добычи, как удильщик; требовать территорию через патруль; общаться и находить себе пару, а также отвлекать или временно ослеплять хищников, чтобы они сбежали. Кроме того, в мезопелагической зоне, куда все еще проникает свет, некоторые организмы маскируются от хищников под ними, освещая свои животы, чтобы соответствовать цвету и интенсивности света сверху, чтобы не было отбрасывания тени. Эта тактика известна как встречное освещение.[12]
Жизненный цикл глубоководных рыб может быть исключительно глубоководным, хотя некоторые виды рождаются на более мелкой воде и тонут по мере созревания. Независимо от глубины, на которой обитают яйца и личинки, они обычно пелагические. Этот планктонный дрейфующий образ жизни требует нейтральной плавучести. Чтобы поддерживать это, яйца и личинки часто содержат в плазме капли масла.[13] Когда эти организмы находятся в полностью созревшем состоянии, они нуждаются в других адаптациях для сохранения своего положения в толще воды. В общем, плотность воды вызывает подъем - аспект плавучести, который заставляет организмы плавать. Чтобы противодействовать этому, плотность организма должна быть больше, чем плотность окружающей воды. Большинство тканей животных плотнее воды, поэтому они должны найти равновесие, чтобы заставить их плавать.[14] Многие организмы развивают плавательные пузыри (газовые полости), чтобы оставаться на плаву, но из-за высокого давления окружающей среды глубоководные рыбы обычно не имеют этого органа. Вместо этого они имеют конструкции, похожие на суда на подводных крыльях, для обеспечения гидродинамической подъемной силы. Также было обнаружено, что чем глубже живет рыба, тем более желеобразной ее плоть и тем более минималистична ее костная структура. Они снижают плотность своей ткани за счет высокого содержания жира, уменьшения веса скелета - достигается за счет уменьшения размера, толщины и содержания минералов - и накопления воды [15] делает их медленнее и менее подвижными, чем поверхностные рыбы.
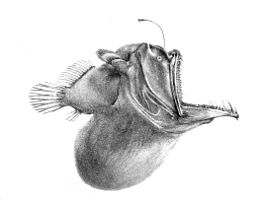
Из-за низкого уровня фотосинтетический свет достигает глубоководной среды, большинству рыб нужно полагаться на органический материя опускается с более высоких уровней или, в редких случаях, гидротермальные источники для питательных веществ. Это делает глубоководные воды намного беднее продуктивность чем более мелкие регионы. Кроме того, животные в пелагической среде немногочисленны, и пища приходит нечасто. Из-за этого организмы нуждаются в адаптации, позволяющей им выжить. У некоторых есть длинные щупальца, которые помогают им находить добычу или привлекать партнеров в кромешной тьме глубокого океана. В частности, у глубоководной рыбы-удильщика на морде выступает длинное приспособление, напоминающее удочку, на конце которого находится биолюминесцентный кусок кожи, который извивается, как червяк, чтобы заманить добычу. Некоторым приходится употреблять в пищу другую рыбу того же размера или больше, чем они, и им нужны приспособления, которые помогут их эффективно переваривать. Большие острые зубы, шарнирные челюсти, непропорционально большой рот и расширяющееся тело - вот некоторые из характеристик, которыми обладают глубоководные рыбы для этой цели.[11] В угорь является одним из примеров организма, который демонстрирует эти характеристики.
Рыбы в различных пелагических и глубоководных бентических зонах имеют физическое строение и поведение, которое заметно отличается друг от друга. Группы сосуществующих видов в каждой зоне, похоже, действуют одинаковым образом, например, небольшой мезопелагический вертикально мигрирующий планктоноеды, батипелагические удильщик, и глубоководные бентосные гремучие хвостики. "[16]
Рэй с плавниками виды с колючими плавниками редки среди глубоководных рыб, что говорит о том, что глубоководные рыбы являются древними и настолько хорошо адаптированы к окружающей среде, что вторжения более современных рыб оказались безуспешными.[17] Те немногие лучевые плавники, которые действительно существуют, в основном находятся в Beryciformes и Lampriformes, которые также являются древними формами. Большинство глубоководных пелагических рыб принадлежат к собственным отрядам, что предполагает длительную эволюцию в глубоководных условиях. Напротив, глубоководные бентосные виды находятся в отрядах, включающих многих родственных мелководных рыб.[18]
| Виды по пелагиали | |
|---|---|
| Многие виды ежедневно перемещаются между зонами вертикальными миграциями. В этой таблице они перечислены в средней или более глубокой зоне, где они регулярно встречаются. | |
| Зона | Виды и видовые группы включают ... |
| Эпипелагический[19] | |
| Мезопелагический | Фонарь, опа, длинноносый ланцет, ячмень, риджхед, саблезуб, свободный стоп-сигнал, морской топорик[20] |
| Батипелагический | В основном щетинистый рот и удильщик. Также клыкастый зуб, гадюка, черный глотатель, телескопрыба, молоток, кинжал, барракудина, черные ножны, бобтейл бекас угорь, единорог, пеликан угорь, дряблый кит. |
| Бентопелагический[19] | Крысиный хвост и Brotula особенно многочисленны. |
| Бентосный | Камбала, миксина, бельдюга, зеленый глаз угорь, скат, пинагой, и летучая мышь[19] |
| Сравнительный состав пелагических рыб | ||||
|---|---|---|---|---|
| Эпипелагический | Мезопелагический | Батипелагический | глубокое море бентосный | |
| мышцы | мускулистые тела, окостеневшие кости, чешуя, хорошо развитые жабры и центральная нервная система, а также большие сердца и почки. | слабо развитый, дряблый | ||
| скелет | крепкие окостеневшие кости | слабая, минимальная оссификация | ||
| напольные весы | да | никто | ||
| нервная система | хорошо развитый | только боковая линия и обоняние | ||
| глаза | большой и чувствительный | маленький и может не работать | переменная (от хорошо развитой до отсутствующей) | |
| фотофоры | отсутствует | общий | общий | обычно отсутствует |
| жабры | хорошо развитый | |||
| почки | большой | маленький | ||
| сердце | большой | маленький | ||
| плавательный пузырь | у вертикально мигрирующих рыб есть плавательный пузырь | сокращен или отсутствует | переменная (от хорошо развитой до отсутствующей) | |
| размер | обычно меньше 25 см | переменная, виды крупнее одного метра не редкость | ||
Мезопелагическая рыба





Ниже эпипелагиали условия быстро меняются. На расстоянии от 200 метров до 1000 метров свет продолжает тускнеть, пока почти не исчезает. Температура падает через термоклин до температуры от 3,9 ° C (39 ° F) до 7,8 ° C (46 ° F). Это сумерки или мезопелагический зона. Давление продолжает расти со скоростью одна атмосфера на каждые 10 метров, в то время как концентрации питательных веществ падают вместе с растворенным кислородом и скоростью, с которой циркулирует вода ».[4]
Операторы гидролокатора, использующие недавно разработанную технологию гидролокатора во время Второй мировой войны, были озадачены тем, что казалось ложным морским дном глубиной 300–500 метров днем и менее глубоким ночью. Оказалось, что это произошло из-за миллионов морских организмов, в первую очередь мелких мезопелагических рыб, плавательные пузыри которых отражают сонар. В сумерках эти организмы мигрируют на мелководье, чтобы питаться планктоном. Слой глубже, когда нет луны, и может становиться мельче, когда над луной проходят облака. Это явление стало известно как глубокий рассеивающий слой.[24]
Большинство мезопелагических рыб производят ежедневно вертикальные миграции, перемещаясь ночью в эпипелагиали, часто следуя аналогичным миграциям зоопланктона, а днем возвращаясь в глубину в целях безопасности.[4][25] Эти вертикальные миграции часто происходят на большие вертикальные расстояния и осуществляются с помощью плавательный пузырь. Плавательный пузырь надувается, когда рыба хочет подняться, и, учитывая высокое давление в мессоплегической зоне, это требует значительных затрат энергии. По мере того, как рыба поднимается, давление в плавательном пузыре должно регулироваться, чтобы он не лопнул. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается.[26] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин, где температура колеблется от 50 ° F (10 ° C) до 69 ° F (20 ° C), таким образом демонстрируя значительные допуски на изменение температуры.[27]
У этих рыб есть мускулистые тела, окостеневшие кости, чешуя, хорошо развитые жабры и центральная нервная система, а также большие сердца и почки. Мезопелагический кормушки планктона иметь маленькие рты с прекрасным жаберные тычинки, в то время рыбоядные имеют более крупный рот и более грубые жаберные тычинки.[4] У вертикально мигрирующих рыб плавательные пузыри.[17]
Мезопелагические рыбы приспособлены к активной жизни в условиях низкой освещенности. Большинство из них - зрительные хищники с большими глазами. У некоторых более глубоководных рыб глаза трубчатые с большими линзами и только стержневые клетки которые смотрят вверх. Они обеспечивают бинокулярное зрение и большую чувствительность к слабым световым сигналам.[4] Эта адаптация обеспечивает улучшенное терминальное зрение за счет бокового зрения и позволяет хищнику различать Кальмар, каракатица, и более мелкие рыбы, которые вырисовываются на фоне мрака над ними.
Мезопелагические рыбы обычно не имеют защитных шипов и используют цвет, чтобы камуфляж сами от других рыб. Засадные хищники темные, черные или красные. Поскольку более длинные красные волны света не достигают глубины моря, красный эффективно действует так же, как черный. Использование миграционных форм затененный серебристые цвета. На животе они часто показывают фотофоры производит низкокачественный свет. Для хищника снизу, смотрящего вверх, это биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют (недостаточный красный) окружающий свет, оставляя биолюминесценцию видимой.[28]
В коричневая рыба-призрак, разновидность ячмень, - единственное известное позвоночное животное, которое использует зеркало вместо линзы для фокусировки изображения в глазах.[29][30]
Отбор проб глубоким траление указывает, что рыба-фонарь составляют до 65% всей глубоководной рыбы биомасса.[31] В самом деле, фонарики являются одними из самых распространенных, густонаселенных и разнообразных из всех. позвоночные, играя важную экологический роль добычи для более крупных организмов. По оценкам, глобальная биомасса фонаря составляет 550-660 миллионов человек. метрические тонны, в несколько раз выловили весь мировой промысел. Фонарь также составляет большую часть биомассы, ответственной за глубокий рассеивающий слой Мирового океана. Сонар отражается от миллионов фонарных рыб плавательные пузыри, создавая видимость ложного дна.[32]
Большеглазый тунец являются эпипелагическим / мезопелагическим видом, питающимся другой рыбой. Спутниковое мечение показало, что большеглазый тунец часто проводит длительные периоды, путешествуя глубоко под поверхностью в дневное время, иногда ныряя на глубину до 500 метров. Считается, что эти движения являются ответом на вертикальные миграции организмов-жертв в глубокий рассеивающий слой.

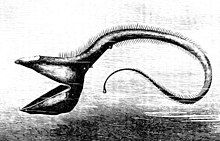
В свободный стоп-сигнал имеет нижняя челюсть четверть длины его тела. Челюсть не имеет пола и крепится только с помощью шарнира и модифицированной кости языка. Крупные клыковидные зубы спереди сменяются множеством мелких зубцов с зазубринами.[33][34]

В свободный стоп-сигнал также одна из немногих рыб, производящих красный биолюминесценция. Поскольку большая часть их добычи не может воспринимать красный свет, это позволяет им охотиться с помощью практически невидимого луча света.[33]

Длиннорылый ланцет. Lancetfish - хищники из засад, которые проводят все свое время в мезопелагической зоне. Это одни из самых крупных мезопелагических рыб (до 2 метров).[35]



Батипелагическая рыба



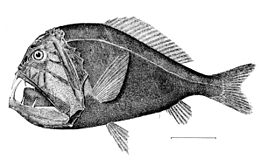
Ниже мезопелагиали кромешная тьма. Это полночь (или батипелагическая зона ), простирающаяся от 1000 метров до дна глубокой воды. бентосная зона. Если вода исключительно глубокая, пелагиали ниже 4000 метров иногда называют нижней полуночью (или абиссопелагическая зона ). Температура в этой зоне колеблется от 1 до 4 градусов по Цельсию и полностью афотична.
Условия в этих зонах несколько однородны; полная темнота, давление сокрушительное, а температура, питательные вещества и уровень растворенного кислорода низкие.[4]
У батипелагических рыб есть особые приспособления чтобы справиться с этими условиями - у них есть медленные метаболизм и неспециализированные диеты, желание есть все, что попадается под руку. Они предпочитают сидеть и ждать еды, а не тратить энергию на ее поиски. Поведение батипелагических рыб можно противопоставить поведению мезопелагических рыб. Мезопелагические рыбы часто очень подвижны, тогда как батипелагические почти все хищники поджидают нас, обычно тратя мало энергии на движение.[44]
Доминирующие батипелагические рыбы - мелкие. щетинистый рот и удильщик; клыкастый зуб, гадюка, кинжал и барракудина также распространены. Эти рыбки маленькие, многие около 10 сантиметров в длину и немногие длиннее 25 см. Они проводят большую часть своего времени, терпеливо ожидая в толще воды, пока не появится добыча или не соблазнится их люминофором. То немногое энергии, которое доступно в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и случайных беспозвоночных или мезопелагических рыб.[44] Около 20 процентов пищи, происходящей из эпипелагиали, приходится на мезопелагиали,[24] но только около 5 процентов попадает в батипелагическую зону.[37]
Батипелагические рыбы ведут оседлый образ жизни, приспособлены к выработке минимальной энергии в среде обитания с очень небольшим количеством пищи или доступной энергии, даже без солнечного света, только с биолюминесценцией. Их тела удлиненный со слабыми водянистыми мышцами и скелетный конструкции. Так как большая часть рыбы состоит из воды, они не сжимаются на этих глубинах. Часто имеют раздвижные, навесные челюсти с загнутыми зубами. Они слизистые, без напольные весы. Центральная нервная система ограничена боковой линией и обонятельной системой, глаза маленькие и могут не функционировать, и жабры, почки и сердце, и плавательные пузыри маленькие или отсутствуют.[37][45]
Это те же особенности, что и у рыб личинки, что предполагает, что в процессе эволюции батипелагические рыбы приобрели эти особенности благодаря неотения. Как и в случае с личинками, эти особенности позволяют рыбе оставаться во взвешенном состоянии в воде с небольшими затратами энергии.[46]
Несмотря на свой свирепый вид, эти глубинные твари в основном представляют собой миниатюрные рыбы со слабыми мускулами и слишком малы, чтобы представлять какую-либо угрозу для людей.
Плавательные пузыри глубоководных рыб либо отсутствуют, либо практически не функционируют, а батипелагические рыбы обычно не совершают вертикальных миграций. Заполнение баллонов при таком большом давлении требует огромных затрат энергии. У некоторых глубоководных рыб есть плавательные пузыри, которые функционируют в молодом возрасте и населяют верхнюю эпипелагическую зону, но они увядают или наполняются жиром, когда рыба перемещается в свою взрослую среду обитания.[47]
Наиболее важными сенсорными системами обычно являются внутреннее ухо, который реагирует на звук, а боковая линия, который реагирует на изменение давления воды. В обонятельный Система также может быть важна для мужчин, которые находят самок по запаху.[48]Батипелагические рыбы черные, а иногда и красные, с небольшими фотофоры. Когда фотофоры используются, они обычно используются для того, чтобы заманить добычу или привлечь партнера. Из-за нехватки пищи батипелагические хищники не избирательны в своих привычках кормления, а хватают все, что подойдет достаточно близко. Они достигают этого за счет большого рта с острыми зубами, чтобы хватать крупную добычу и перекрывать друг друга. жаберные тычинки которые не позволяют маленькой добыче, проглоченной, ускользнуть.[45]
Найти себе пару в этой зоне непросто. Некоторые виды зависят от биолюминесценция, где биолюминесцентные узоры уникальны для конкретных видов. Другие гермафродиты, что удваивает их шансы произвести яйцеклетки и сперму при встрече.[37] Самка удильщика выпускает феромоны для привлечения крошечных самцов. Когда самец находит ее, он кусает ее и никогда не отпускает. Когда самец удильщиков Гаплофрин моллис впивается в кожу самки, он выпускает фермент который переваривает кожу его рта и ее тело, сплавляя пару до точки соединения двух кровеносных систем. Затем самец атрофируется, превращаясь в пару гонады. Этот крайний половой диморфизм гарантирует, что когда самка будет готова к нересту, у нее сразу же появится помощник.[49]
В батипелагической зоне обитают многие другие формы, такие как кальмары, большие киты, осьминоги, губки, брахиоподы, морские звезды и ехиноиды, но в этой зоне рыбам жить сложно.

В пеликан угорь использует свой рот как сеть, открывая большой рот и плывя к своей добыче. На конце хвоста у него есть люминесцентный орган, привлекающий добычу.

В черный глотатель, с расширяющимся желудком, отличается способностью глотать целиком, костлявые рыбы в десять раз больше его массы.[50][51]

женский Гаплофрин моллис удильщик с прикрепленными самцами, которые атрофировались в пару гонады, для использования, когда самка готова к нересту.


Адаптация к высокому давлению
По мере того, как рыба продвигается все глубже в море, вес воды над ее головой увеличивается. гидростатическое давление на рыбе. Это повышенное давление составляет примерно один стандартная атмосфера на каждые 10 метров глубины (одна атмосфера - это давление, оказываемое на поверхность моря атмосферой выше). Для рыбы на дне батипелагическая зона, это давление составляет около 400 атмосфер (около 6000 фунтов на квадратный дюйм).[52]
Глубоководные организмы обладают адаптациями на клеточном и физиологическом уровнях, которые позволяют им выживать в условиях сильного давления. Отсутствие этих приспособлений ограничивает глубины, на которых могут существовать мелководные виды. Высокий уровень внешнего давления влияет на протекание метаболических процессов и биохимических реакций.Равновесие многих химических реакций нарушается давлением, и давление может тормозить процессы, которые приводят к увеличению объема. Вода, которая играет ключевую роль во многих биологических процессах, очень чувствительна к изменениям объема, главным образом потому, что компоненты клеточной жидкости влияют на структуру воды. Таким образом, ферментативные реакции, вызывающие изменения в организации воды, эффективно изменяют объем системы.[53] Белки, ответственные за катализирующие реакции, обычно удерживаются вместе слабыми облигации и реакции обычно включают увеличение объема.[54] Чтобы адаптироваться к этому изменению, структура белка и критерии реакции глубоководных рыб были адаптированы, чтобы выдерживать давление для проведения реакций в этих условиях. В средах высокого давления двухслойный клеточные мембраны испытывают потерю текучести. Глубоководные клеточные мембраны благоприятствуют фосфолипидные бислои с более высокой долей ненасыщенные жирные кислоты, которые обладают большей текучестью, чем их аналоги на уровне моря.

Глубоководные виды демонстрируют меньшие изменения энтропия и энтальпия по сравнению с организмами на уровне поверхности, поскольку окружающая среда с высоким давлением и низкой температурой способствует отрицательным изменениям энтальпии и снижает зависимость от реакций, вызванных энтропией. Со структурной точки зрения, глобулярные белки глубоководной рыбы третичная структура G-актин относительно жесткий по сравнению с рыбой, расположенной на поверхности.[55] То, что белки глубоководных рыб структурно отличаются от поверхностных рыб, очевидно из наблюдения, что актин из мышечные волокна глубоководных рыб чрезвычайно жаропрочные; аналогично тому, что находится в ящерицы. Эти белки структурно усилены за счет модификации связей в третичной структуре белка, что также вызывает высокие уровни термической стабильности.[56] Белки структурно усилены, чтобы противостоять давлению за счет модификации связей в третичной структуре.[57] Следовательно, высокий уровень гидростатического давления, аналогичный высоким температурам тела термофильных пустынных рептилий, способствует жестким белковым структурам.
Na + / K + -АТФаза это липопротеин фермент, который играет важную роль в осморегуляция и сильно зависит от гидростатического давления. Ингибирование Na + / K + -АТФазы происходит из-за увеличения сжатия из-за давления. Ограничивающая скорость стадия реакции Na + / K + -АТФазы вызывает расширение бислоя, окружающего белок, и, следовательно, увеличение объема. Увеличение объема делает реактивность Na + / K + -АТФазы чувствительной к более высоким давлениям. Несмотря на то, что активность Na + / K + -АТФазы на грамм жаберная ткань ниже у глубоководных рыб, Na + / K + -АТФазы глубоководных рыб демонстрируют гораздо более высокую устойчивость к гидростатическому давлению по сравнению с их мелководными аналогами. Это проиллюстрировано между видами С. акролепис (глубина около 2000 м) и его хадальпелагический двойник C. armatus (глубина около 4000 м), где Na + / K + -АТФазы C. armatus гораздо менее чувствительны к давлению. Это сопротивление давлению можно объяснить адаптацией белков и липидов. части Na + / K + -АТФазы.[58]
Фонарь

Отбор проб глубоким траление указывает, что рыба-фонарь составляют до 65% всей глубоководной рыбы биомасса.[31] В самом деле, фонарики являются одними из самых распространенных, густонаселенных и разнообразных из всех. позвоночные, играя важную экологический роль добычи для более крупных организмов. По оценкам, мировая биомасса составляет 550-660 миллионов человек. метрических тонн, в несколько раз больше улова мирового рыболовства, фонарь также составляет большую часть биомассы, ответственной за глубокий рассеивающий слой Мирового океана. в Южный океан, Миктофиды являются альтернативным пищевым ресурсом для криль для хищников, таких как Кальмар и королевский пингвин. Хотя эти рыбы многочисленны и плодовиты, в настоящее время только несколько промышленных фонарей. рыболовство Существуют: Сюда входят ограниченные операции Южная Африка, в субантарктический, а в Оманский залив.
Вымирающие виды
2006 г. изучение от Канадский ученые нашел пять видов глубоководных рыб - синий хек, колючий угорь - оказаться на грани исчезновения из-за смещения коммерческое рыболовство от континентальные шельфы на склоны континентальные шельфы, до глубины 1600 метров. Медленный воспроизведение из этих рыбы - они достигают половая зрелость примерно в том же возрасте, что и люди - одна из основных причин того, что они не могут оправиться от чрезмерная рыбалка.[59]
Смотрите также
- Перепись морской жизни
- Глубокая океанская вода
- Глубокое море
- Глубоководные сообщества
- Глубоководная рыба
- Демерсальная рыба
- Пелагическая рыба
Цитаты
- ^ Трухильо, Алан П .; Гарольд В. Турман (2011). Основы океанографии 10-е изд.. Бостон: Прентис Холл. п. 354. ISBN 978-0321668127.
- ^ Трухильо, Алан П .; Гарольд В. Турман (2011). Основы океанографии 10-е изд.. Бостон: Прентис Холл. п. 365. ISBN 978-0321668127.
- ^ Трухильо, Алан П .; Гарольд В. Турман (2011). Основы океанографии 10-е изд.. Бостон: Прентис Холл. С. 457, 460. ISBN 978-0321668127.
- ^ а б c d е ж Мойл и Чех, 2004 г., стр. 585
- ^ Wharton, Дэвид. (2002). Жизнь на пределе возможностей: организмы в экстремальных условиях. Кембридж, Великобритания: Cambridge UP. стр.198. ISBN 978-0521782128.
- ^ Уортон, Дэвид (2002). Жизнь на пределе возможностей: организмы в экстремальных условиях. Кембридж, Великобритания: Cambridge UP. С. 199, 201–202. ISBN 978-0521782128.
- ^ а б Wharton, Дэвид. (2002). Жизнь на пределе возможностей: организмы в экстремальных условиях. Кембридж, Великобритания: Cambridge UP. стр.199. ISBN 978-0521782128.
- ^ Compagno, L.J.V. (1984). Акулы мира: аннотированный и иллюстрированный каталог известных на сегодняшний день видов акул. Продовольственная и сельскохозяйственная организация Объединенных Наций. С. 14–15. ISBN 92-5-101384-5.
- ^ Уортон, Дэвид А. (2007-07-23). Жизнь на пределе возможностей: организмы в экстремальных условиях. Издательство Кембриджского университета. ISBN 9781139431941.
- ^ Мусилова, Зузана; Кортези, Фабио; Матчинер, Майкл; Дэвис, Уэйн; Патель, Джагдиш; Стиб, Сара; де Буссероль, Фанни; Мальмстрём, Мартин; Торресен, Оле; Браун, Селеста; Маунтфорд, Джессика; Ханель, Рейнхольд; Стенкамп, Дебора; Якобсен, Кьетилль; Карлтон, Карен; Джентофт, Сиссель; Маршалл, Джастин; Зальцбургер, Вальтер. «Видение с использованием нескольких различных стержневых опсинов у глубоководных рыб». Sciencemag. Американская ассоциация развития науки. Получено 10 мая 2019.
- ^ а б Трухильо, Алан П .; Гарольд В. Турман (2011). Основы океанографии 10-е изд.. Бостон: Прентис Холл. п. 415. ISBN 978-0321668127.
- ^ Трухильо, Алан П .; Гарольд В. Турман (2011). Основы океанографии 10-е изд.. Бостон: Прентис Холл. С. 414–415. ISBN 978-0321668127.
- ^ Рэндалл, Дэвид Дж .; Энтони Питер Фаррелл (1997). Глубоководные рыбы. Сан-Диего: академический. п. 217. ISBN 978-0123504401.
- ^ Рэндалл, Дэвид Дж .; Энтони Питер Фаррелл (1997). Глубоководные рыбы. Сан-Диего: академический. п. 195. ISBN 978-0123504401.
- ^ Рэндалл, Дэвид Дж .; Энтони Питер Фаррелл (1997). Глубоководные рыбы. Сан-Диего: академический. С. 196, 225. ISBN 978-0123504401.
- ^ Мойл и Чех, 2004, с. 591
- ^ а б Хаэдрих Р.Л. (1996) «Глубоководные рыбы: эволюция и адаптация в крупнейших жизненных пространствах Земли» Журнал биологии рыб 49(SA): 40-53.
- ^ Мойл и Чех, 2004 г., стр. 586
- ^ а б c Мойл и Чех, 2004 г., стр. 571
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Argyropelecus aculeatus" в FishBase. Версия от августа 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Dissostichus mawsoni" в FishBase. Версия от августа 2009 г.
- ^ Раскрыта тайна глубоководной рыбы с трубчатыми глазами и прозрачной головой ScienceDaily, 24 февраля 2009 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Гигантура чуни" в FishBase. Версия от октября 2010 г.
- ^ а б Райан П. «Глубоководные существа: мезопелагическая зона» Те Ара - Энциклопедия Новой Зеландии. Обновлено 21 сентября 2007 г.
- ^ Bone & Moore 2008, стр. 38.
- ^ Дуглас Е.Л., Фридл В.А., Пиквелл Г.В. (1976). «Рыбы в зонах минимума кислорода: характеристики оксигенации крови». Наука. 191 (4230): 957–959. Дои:10.1126 / science.1251208. PMID 1251208.
- ^ Мойл и Чех, 2004, с. 590
- ^ Munz WRA (1976). «О желтых линзах у мезопелагических животных». Морская биологическая ассоциация Великобритании. 56 (4): 963–976. Дои:10.1017 / S0025315400021019.
- ^ Wagner, HJ; Douglas, R.H .; Франк, T.M .; Робертс, Н.В. И Партридж, Дж. К. (27 января 2009 г.). «Новый глаз позвоночных животных, использующий как рефракционную, так и отражающую оптику». Текущая биология. 19 (2): 108–114. Дои:10.1016 / j.cub.2008.11.061. PMID 19110427. S2CID 18680315.
- ^ Смит, Л. (8 января 2009 г.). «Рыба с четырьмя глазами может видеть сквозь мрак глубокого моря». Times Online. Times Newspapers Ltd. Проверено 14 марта 2009 г.
- ^ а б Халли, П. Александр (1998). Paxton, J.R .; Эшмайер, W.N. (ред.). Энциклопедия рыб. Сан-Диего: Academic Press. С. 127–128. ISBN 0-12-547665-5.
- ^ Р. Корнехо; Р. Коппельманн; Т. Саттон. «Разнообразие глубоководных рыб и экология в придонном пограничном слое».
- ^ а б Kenaley, C.P (2007). "Пересмотр семейства Stoplight Loosejaw" Малакостей (Teleostei: Stomiidae: Malacosteinae), с описанием нового вида из умеренного южного полушария и Индийского океана ». Копея. 2007 (4): 886–900. Дои:10.1643 / 0045-8511 (2007) 7 [886: ROTSLG] 2.0.CO; 2.
- ^ Саттон, Т.Т. (ноябрь 2005 г.). «Трофическая экология глубоководных рыб. Malacosteus niger (Рыбы: Stomiidae): Загадочная экология кормления, обеспечивающая уникальную зрительную систему? ". Deep-Sea Research Part I: Oceanographic Research Papers. 52 (11): 2065–2076. Дои:10.1016 / j.dsr.2005.06.011.
- ^ Мойл и Чех, 2004, с. 336
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Anotopterus pharao" в FishBase. Версия от апреля 2010 г.
- ^ а б c d Райан П. «Глубоководные существа: Батипелагическая зона» Те Ара - Энциклопедия Новой Зеландии. Обновлено 21 сентября 2007 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2006). "Гоностома батифильная" в FishBase. Версия от января 2006 г.
- ^ Froese, Rainer и Daniel Pauly, ред. (2009). «Гоностома» в FishBase. Версия от августа 2009 г.
- ^ «Объединяя знания и людей более 10 лет». Архивировано из оригинал на 2012-07-09.
- ^ «Ученые разгадывают тайну: все 3 рыбы одинаковые». 22 января 2009 г.. Получено 2009-01-22.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2010). "Chauliodus sloani" в FishBase. Версия от апреля 2010 г.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Anoplogaster cornuta" в FishBase. Версия от августа 2009 г.
- ^ а б Мойл и Чех, 2004, с. 594
- ^ а б Мойл и Чех, 2004, с. 587
- ^ Маршалл (1984) "Прогенетические тенденции у глубоководных рыб", стр. 91-101 в Potts GW и Wootton RJ (ред.) (1984) Размножение рыб: стратегии и тактика Общество рыболовства Британских островов.
- ^ Хорн MH (1970). «Плавательный пузырь как ювенильный орган строматеидных рыб». Breviora. 359: 1–9.
- ^ Прыгун GY, Bair RC (1991). "Местоположение по обонянию: модель и приложение к проблеме спаривания у глубоководных топориков. Argyropelecus hemigymnus". Американский натуралист. 138 (6): 1431–1458. Дои:10.1086/285295. JSTOR 2462555. S2CID 84386858.
- ^ Теодор В. Питч (1975). «Преждевременный сексуальный паразитизм в глубоководных цератиоидных удильщиках, Cryptopsaras couesi Gill». Природа. 256 (5512): 38–40. Дои:10.1038 / 256038a0. S2CID 4226567. Получено 31 июля 2008.
- ^ Джордан, Д.С. (1905). Руководство по изучению рыб. Х. Холт и компания.
- ^ Froese, Rainer and Pauly, Daniel, eds. (2009). "Chiasmodon niger" в FishBase. Версия от августа 2009 г.
- ^ Скотт, Томас Р .; Пауэлл, Джеймс (2018-09-24). Вселенная, как она есть на самом деле: Земля, космос, материя и время. Издательство Колумбийского университета. Дои:10.7312 / scot18494. ISBN 978-0-231-54576-1.
- ^ ХОЧАЧКА, ПЕТР ЗАНЯТИЕ СОМЕРО, ДЖОРДЖ Н. (1984). Биохимическая адаптация. Издательство Принстонского университета. JSTOR j.ctt7zv9d4.
- ^ Приеде, Имантс Г., изд. (2017), «Адаптации к глубокому морю», Глубоководные рыбы: биология, разнообразие, экология и рыболовство, Кембридж: Издательство Кембриджского университета, стр. 87–138, ISBN 978-1-316-01833-0, получено 2020-10-31
- ^ ХОЧАЧКА, ПЕТР ЗАНЯТОЙ СОМЕРО, ДЖОРДЖ Н. (1984). Биохимическая адаптация. Издательство Принстонского университета. JSTOR j.ctt7zv9d4.
- ^ Приеде, Имантс Г., изд. (2017), «Адаптации к глубокому морю», Глубоководные рыбы: биология, разнообразие, экология и рыболовство, Кембридж: Издательство Кембриджского университета, стр. 87–138, ISBN 978-1-316-01833-0, получено 2020-10-31
- ^ Приеде, Имантс Г., изд. (2017), «Адаптации к глубокому морю», Глубоководные рыбы: биология, разнообразие, экология и рыболовство, Кембридж: Издательство Кембриджского университета, стр. 87–138, ISBN 978-1-316-01833-0, получено 2020-10-31
- ^ Somero, G. N (октябрь 1992 г.). «Адаптация к высокому гидростатическому давлению». Ежегодный обзор физиологии. 54 (1): 557–577. Дои:10.1146 / annurev.ph.54.030192.003013. ISSN 0066-4278. PMID 1314046.
- ^ Дивайн Дженнифер А., Бейкер Криста Д., Хедрих Ричард Л. (2006). «Рыболовство: глубоководные рыбы считаются исчезающими». Природа. 439 (7072): 29. Дои:10.1038 / 439029a. PMID 16397489. S2CID 4428618.CS1 maint: несколько имен: список авторов (ссылка на сайт)
использованная литература
- Мойл, ПБ, и Чех, Джей Джей (2004) Рыбы, Введение в ихтиологию. 5-е изд., Бенджамин Каммингс. ISBN 978-0-13-100847-2
дальнейшее чтение
- Гордон Дж. Д. М. (2001) «Глубоководные рыбы» В: Джон Х. Стил, Стив А. Торп, Карл К. Турекиан (ред.) Элементы физической океанографии, страницы 227–233, Academic Press. ISBN 9780123757241.
- Хоар WS, Рэндалл DJ и Фаррелл AP (редакторы) (1997) Глубоководные рыбы, Academic Press. ISBN 9780080585406.
- Шоттон, Росс (1995) «Глубоководное рыболовство» В: Обзор состояния мировых морских рыбных ресурсов, Технический документ ФАО по рыболовству 457, ФАО, Рим. ISBN 92-5-105267-0.
- Тандстад М., Шоттон Р., Сандерс Дж. И Кароччи Ф. (2011) «Глубоководное рыболовство» В: Обзор состояния мировых морских рыбных ресурсов, страницы 265–278, Технический документ ФАО по рыболовству 569, ФАО, Рим. ISBN 978-92-5-107023-9.
внешние ссылки
| Внешнее видео | |
|---|---|
- https://www.pbs.org/wgbh/nova/abyss/life/bestiary.html
- http://ocean.nationalgeographic.com/ocean/photos/deep-sea-creatures/
- Глубоководные существа - Статьи, факты и изображения глубоководных животных


| Около рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| От место обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||
| Авторитетный контроль |
|---|