Криль - Krill
| Криль | |
|---|---|
 | |
| Северный криль (Meganyctiphanes norvegica) | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Подтип: | Ракообразные |
| Учебный класс: | Малакострака |
| Суперзаказ: | Эукарида |
| Заказ: | Euphausiacea Дана, 1852 |
| Семьи и роды | |
| |
Криль маленькие ракообразные из порядок Euphausiacea, и встречаются во всех океанах мира. Название «криль» происходит от норвежский язык слово криль, что означает "маленький жарить рыбы »,[1] который также часто относят к видам рыб.
Криль считается важным трофический уровень подключение - в нижней части пищевая цепочка. Они питаются фитопланктон и (в меньшей степени) зоопланктон, но также являются основным источником пищи для многих крупных животных. в Южный океан, один вид, Антарктический криль, Euphausia superba, составляет оценочную биомасса около 379 000 000 тонн,[2] что делает его одним из видов с наибольшей общей биомассой. Более половины этой биомассы ежегодно поедается китами, тюленями, пингвинами, кальмарами и рыбой. Большинство видов криля имеют большие размеры. ежедневные вертикальные миграции, таким образом обеспечивая пищу хищникам у поверхности ночью и в более глубоких водах днем.
Коммерческий промысел криля ведется в Южном океане и в водах вокруг Японии. Общий мировой урожай составляет 150 000–200 000 тонн в год, большая часть из которых приходится на Море Скотия. Большая часть улова криля используется для аквакультура и аквариум кормит, как приманка в спортивная рыбалка, или в фармацевтической промышленности. В Японии, на Филиппинах и в России криль также используется в пищу человеком и известен как окиами (オ キ ア ミ) в Японии. Их едят как камаронес в Испании и на Филиппинах. На Филиппинах криль также известен как Аламанг и используются для приготовления соленой пасты, называемой багун.
Криль также является основной добычей усатые киты, в том числе синий кит.
Таксономия
Криль принадлежит к большим членистоногие подтип, то Ракообразные. Самая известная и самая большая группа ракообразных, учебный класс Малакострака, включает суперзаказ Эукарида состоящие из трех отрядов: Euphausiacea (криль), Декапода (креветки, креветки, лобстеры, крабы) и планктонные Амфионидацея.
Отряд Euphausiacea состоит из двух семьи. Более обильный Euphausiidae содержит 10 различных роды всего 85 видов. Из них род Евфаузия является самым крупным, насчитывает 31 вид.[3] Менее известная семья, Bentheuphausiidae, имеет только один разновидность, Бентуфаузия амблиопс, а батипелагический криль, обитающий на глубине ниже 1000 м (3300 футов). Считается самым примитивным из сохранившихся видов криля.[4]
Хорошо известные виды Euphausiidae промыслового крилевый промысел включают Антарктический криль (Euphausia superba), Тихоокеанский криль (E. pacifica) и Северный криль (Meganyctiphanes norvegica).[5]
Филогения
| Предлагаемая филогения Euphausiacea[6] | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Филогения, полученная на основе морфологических данных, (names) названия, введенные в,[6] (♣) возможно парафилетический таксон из-за Нематобрахия в.[6] (♦) clades отличается от Casanova (1984),[7] куда Псевдоевфаузия сестра Никтифан, Евфаузия сестра Thysanopoda и Нематобрахия сестра Стилохейрон. |
По состоянию на 2013 год[Обновить], отряд Euphausiacea считается монофилетический из-за нескольких уникальных сохраненных морфологических характеристик (аутапоморфия ), такие как голые нитчатые жабры и тонкие торакоподы.[8] и молекулярными исследованиями.[9][10][11]
Существует много теорий о местонахождении отряда Euphausiacea. Поскольку первое описание Тизанопод трикуспид к Анри Милн-Эдвардс в 1830 г. сходство их двуветвистых торакопод привело зоологов к группировке эвфаузиид и мизидацея в отряд Шизоподы, который был разделен Йохан Эрик Вести Боас в 1883 г. на два отдельных ордена.[12] Потом, Уильям Томас Калман (1904) занял первое место Mysidacea в суперзаказе Peracarida и эвфаузииды в надотряде Эукарида, хотя еще до 1930-х годов отряд Schizopoda был защищен.[8] Позже было также предложено, чтобы отряд Euphausiacea был сгруппирован с Penaeidae (семейство креветок) у Decapoda на основании сходства в развитии, как отмечает Роберт Герни и Изабелла Гордон.[13][14] Причина этой дискуссии заключается в том, что криль имеет некоторые морфологические черты десятиногих и других мизид.[8]
Молекулярные исследования не сгруппировали их однозначно, возможно, из-за нехватки ключевых редких видов, таких как Бентуфаузия амблиопс в криле и Amphionides reynaudii в Эукариде. Одно исследование поддерживает монофилию Eucarida (с базальной Mysida),[15] другие группы Euphausiacea с Mysida (Schizopoda),[10] в то время как еще одна группа Euphausiacea с Hoplocarida.[16]
График
Никакие сохранившиеся ископаемые останки нельзя однозначно отнести к Euphausiacea. Некоторые вымершие Eumalacostracan таксоны считались эвфаузийцами, такими как Антракофаузия, Крангопсис - теперь закреплен за Эшронектида (Hoplocarida)[6] - и Палеомизис.[17] Все датировки видообразование события оценивались молекулярные часы методы, которые поместили последнего общего предка семейства криля Euphausiidae (отряд Euphausiacea minus Бентуфаузия амблиопс) чтобы жить в Нижний мел о 130 миллион лет назад.[10]
Распределение
Криль встречается во всем мире во всех океанах, хотя многие отдельные виды эндемичный или же неритический (т.е. прибрежные) распространения. Бентуфаузия амблиопс, а батипелагический вид, имеет космополитическое распространение в его глубоководной среде обитания.[18]
Виды рода Thysanoessa встречаются в обоих Атлантический и Тихий океан океаны.[19] Тихий океан является домом для Евфаузия тихая. Северный криль встречается через Атлантику от Средиземное море на север.
К видам с неритическим распределением относятся четыре вида рода Никтифан.[20] Их очень много вдоль апвеллинг регионы Калифорния, Гумбольдт, Бенгела, и Канарские острова текущие системы.[21][22][23] Другой вид, имеющий только неритическое распространение, - это E. crystalorophias, который является эндемиком побережья Антарктики.[24]
Виды с эндемичным распространением включают: Nyctiphanes capensis, которое встречается только в Бенгельском течении,[20] E. mucronata в течении Гумбольдта,[25] и шесть Евфаузия родом из Южного океана.
В Антарктике известно семь видов,[26] один в роду Thysanoessa (T. macrura ) и шесть в Евфаузия. В Антарктический криль (Euphausia superba) обычно обитает на глубине до 100 м (330 футов),[27] тогда как ледяной криль (Евфаузия кристалорфиас ) достигают глубины 4000 м (13 100 футов), хотя обычно они обитают на глубинах не более 300–600 м (1000–2000 футов).[28] Оба находятся в широты к югу от 55 ° ю.ш., с E. crystalorophias доминирующий к югу от 74 ° ю.ш.[29] и в регионах паковый лед. Другие виды, известные в Южный океан находятся E. frigida, E. longirostris, E. triacantha и Э. валлентини.[30]
Анатомия и морфология
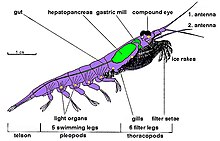

Криль декапод ракообразные и, как и все ракообразные, у них хитиновый внешний каркас. У них есть стандарт анатомия декапода с их телами, состоящими из трех части: головогрудь состоит из голова и грудная клетка которые слиты, плеон который несет десять плавательных ног и хвостовой веер. Эта внешняя оболочка криля прозрачна у большинства видов.
Особенности криля замысловатые сложные глаза. Некоторые виды приспосабливаются к различным условиям освещения с помощью экранирования. пигменты.[31]
У них есть два усики и несколько пар грудных ног, называемых переоподы или же торакоподы, названы так потому, что прикрепляются к грудной клетке. Их количество варьируется в зависимости от рода и вида. Эти грудные ноги включают ноги для кормления и для ухода за ногами.
Криль - это декодоногие моллюски, поэтому у всех видов по пять пар плавательные ноги так называемые «пловцы», очень похожие на Омар или же пресноводные раки.
Большая часть криля имеет длину примерно 1-2 сантиметра (0,4-0,8 дюйма), чем взрослые особи. Некоторые виды вырастают до размеров порядка 6–15 сантиметров (2,4–5,9 дюйма). Самый крупный вид криля, Thysanopoda spinicauda, жизни глубоко в открытом океане.[32] Криль можно легко отличить от других ракообразных, таких как настоящие креветка по их внешнему виду жабры.[33]
Кроме Бентуфаузия амблиопс, криль биолюминесцентный животные, имеющие органы, называемые фотофоры которые могут излучать свет. Свет генерируется фермент -катализированный хемилюминесценция реакция, в которой люциферин (своего рода пигмент) активируется люцифераза фермент. Исследования показывают, что люциферин многих видов криля является флуоресцентный тетрапиррол похож, но не идентичен динофлагеллята люциферин[34] и что криль, вероятно, сам не производит это вещество, а получает его как часть своего рациона, который содержит динофлагеллаты.[35] Фотофоры криля - это сложные органы с линзами и способностями фокусировки, которые могут вращаться мышцами.[36] Точная функция этих органов пока неизвестна; Возможности включают совокупление, социальное взаимодействие или ориентацию, а также в качестве формы маскировки противосветового освещения, чтобы компенсировать их тень от окружающего света над головой.[37][38]
Экология

Кормление
Многие криль питатели-фильтры:[22] их самые передовые придатки торакоподы образуют очень тонкие соты, с помощью которых они могут фильтровать пищу из воды. Эти фильтры могут быть очень тонкими у видов (например, Евфаузия spp.), которые питаются в основном фитопланктон, в частности на диатомеи, которые являются одноклеточными водоросли. Криль в основном всеядный,[40] хотя некоторые виды плотоядный, охотясь на маленькие зоопланктон и рыба личинки.[41]
Криль - важный элемент водного пищевая цепочка. Криль преобразовал основное производство их добычи в форму, пригодную для потребления более крупными животными, которые не могут питаться непосредственно крошечными водорослями. Северный криль и некоторые другие виды имеют относительно небольшую фильтрующую корзину и активно охотятся. копеподы и более крупный зоопланктон.[41]
Хищничество
Крилем питаются многие животные, от более мелких животных, таких как рыбы или же пингвины к большим, как уплотнения и усатые киты.[42]
Нарушения экосистема приведение к сокращению популяции криля может иметь далеко идущие последствия. Во время кокколитофора цвести в Берингово море в 1998 г.,[43] например, концентрация диатомовых водорослей упала в пораженной области. Криль не может питаться более мелкими кокколитофоридами, и, следовательно, популяция криля (в основном E. pacifica) в этом регионе резко сократилось. Это, в свою очередь, повлияло на другие виды: буревестник население упало. Считалось, что инцидент был одной из причин лосось в тот сезон не появлялись.[44]
Несколько одноклеточных эндопаразитоидный инфузории рода Collinia может заразить виды криля и уничтожить затронутые популяции. Сообщалось о таких заболеваниях Thysanoessa inermis в Беринговом море, а также для E. pacifica, Thysanoessa spinifera, и T. gregaria у побережья Тихого океана в Северной Америке.[45][46] Немного эктопаразиты семьи Dajidae (эпикаридий изоподы ) поражают криль (а также креветки и мизиды ); один такой паразит Oculophryxus bicaulis, который был найден на криле Стилохейрон аффинный и С. longicorne. Он прикрепляется к стеблю глаза животного и высасывает кровь из его головы; очевидно, что это препятствует воспроизводству хозяина, поскольку ни одно из пораженных животных не достигло зрелости.[47]
Изменение климата представляет собой еще одну угрозу для популяций криля.[48]
Пластмассы
Предварительные исследования показывают, что криль может переваривать микропластик диаметром менее 5 мм (0,20 дюйма), разрушая их и выводя обратно в окружающую среду в меньшем виде.[49]
История жизни и поведение

Жизненный цикл криля относительно хорошо изучен, несмотря на незначительные различия в деталях от вида к виду.[13][22] После вылупления криля они проходят несколько личиночных стадий:науплий, псевдометанауплиус, метанауплий, калиптопс, и фурцилия, каждый из которых делится на подэтапы. Стадия псевдометанауплиуса присуща только видам, откладывающим яйца в яйцевидном мешочке: так называемые «мешочки-производители». Личинки разрастаются и линька многократно по мере развития, заменяя свой жесткий экзоскелет, когда он становится слишком маленьким. Более мелкие животные линяют чаще, чем более крупные. Желток Запасы внутри их тела питают личинок на стадии метанауплиуса.
По стадиям калиптопса дифференциация прогрессировало достаточно далеко, чтобы у них развились ротовая полость и пищеварительный тракт, и они начали есть фитопланктон. К этому времени их запасы желтка исчерпаны, и личинки должны достичь фотическая зона, верхние слои океана, где процветают водоросли. На стадиях фурцилий добавляются сегменты с парами плавников, начиная с самых передних сегментов. Каждая новая пара становится функциональной только при следующей линьке. Количество сегментов, добавляемых на любой из стадий фурцилии, может варьироваться даже в пределах одного вида в зависимости от условий окружающей среды.[50] После финальной стадии фурцилии из незрелой молоди появляется форма, похожая на взрослую особь, и впоследствии она развивается. гонады и созревает половым путем.[51]
Размножение

Во время брачного сезона, который зависит от вида и климата, самец откладывает мешок для спермы у генитального отверстия самки (названа thelycum). Самки могут нести в своем яичник, который в таком случае может составлять до одной трети массы тела животного.[52] У криля может быть несколько выводков за один сезон, с интервалами между выводками, длящимися порядка дней.[23][53]
Криль использует два типа нерестового механизма.[23] 57 видов родов Бентеуфаузия, Евфаузия, Меганиктифаны, Thysanoessa, и Thysanopoda являются «распространителями размножения»: самка выпускает оплодотворенные икры в воду, где они обычно тонут, рассредоточиваются и остаются сами по себе. Эти виды обычно вылупляются на стадии науплиуса 1, но недавно было обнаружено, что они вылупляются иногда на стадии метанауплиуса или даже на стадии калиптописа.[54] Остальные 29 видов других родов являются «производителями мешочков», когда самка несет с собой яйца, прикрепленные к последним парам торакопод, пока они не вылупятся в виде метанауплиусов, хотя некоторые виды любят Nematoscelis difficilis может вылупляться как науплий или псевдометанауплиус.[55]
Линька
Линька происходит всякий раз, когда особь вырастает из своего жесткого экзоскелета. Молодые животные, растущие быстрее, линяют чаще, чем более старые и крупные. Частота линьки широко варьируется в зависимости от вида и даже в пределах одного вида зависит от многих внешних факторов, таких как широта, температура воды и наличие пищи. Субтропический вид Никтифан симплексНапример, общий период между линькой составляет от двух до семи дней: личинки линяют в среднем каждые четыре дня, а молодые и взрослые особи - в среднем каждые шесть дней. За E. superba в антарктическом море наблюдались периоды между линьками от 9 до 28 дней в зависимости от температуры от -1 до 4 ° C (30 и 39 ° F), а для Meganyctiphanes norvegica в Северное море периоды между линьками также варьируются от 9 до 28 дней, но при температуре от 2,5 до 15 ° C (от 36,5 до 59,0 ° F).[56] E. superba способен уменьшать размер своего тела, когда не хватает пищи, линяет также, когда его экзоскелет становится слишком большим.[57] Подобная усадка также наблюдалась для E. pacifica, вид, обитающий в Тихом океане от полярных до умеренных зон в результате адаптации к аномально высоким температурам воды. Постулируется усадка и для других видов криля умеренной зоны.[58]
Срок жизни
Некоторые высокоширотные виды криля могут жить более шести лет (например, Euphausia superba); другие, такие как среднеширотные виды Евфаузия тихая, проживи всего два года.[5] Субтропический или тропический продолжительность жизни видов еще меньше, например, Никтифан симплекс, который обычно живет всего шесть-восемь месяцев.[59]
Роение
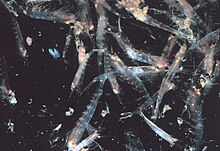
Большинство криля роение животные; размеры и плотность таких стай варьируются в зависимости от вида и региона. За Euphausia superbaстаи достигают от 10 000 до 60 000 особей на кубический метр.[60][61] Рой - это защитный механизм, сбивающий с толку более мелких хищников, которые хотели бы выделить особей. В 2012 году Гандоми и Алави представили то, что кажется успешный стохастический алгоритм для моделирования поведения стай криля. Алгоритм основан на трех основных факторах: «(i) движение, вызванное присутствием других особей, (ii) добыча пищи, и (iii) случайное распространение».[62]
Вертикальная миграция

Криль обычно следует за дневной вертикальная миграция. Было высказано предположение, что они проводят день на большей глубине и ночью поднимаются к поверхности. Чем глубже они заходят, тем меньше их активность,[63] очевидно, чтобы уменьшить количество встреч с хищниками и сберечь энергию. Плавательная активность криля зависит от наполненности желудка. Насытившиеся животные, кормившиеся на поверхности, менее активно плавают и поэтому опускаются ниже смешанного слоя.[64] Когда они тонут, они производят кал что подразумевает роль в Антарктике цикл углерода. Криль с пустым желудком плавает более активно и, таким образом, устремляется к поверхности.
Вертикальная миграция может происходить 2–3 раза в день. Некоторые виды (например, Euphausia superba, E. pacifica, Э. хансени, Pseudeuphausia latifrons, и Thysanoessa spinifera) образуют в течение дня стаи на поверхности для кормления и воспроизводства, хотя такое поведение опасно, поскольку делает их чрезвычайно уязвимыми для хищников.[65]
Экспериментальные исследования с использованием Артемия салина в качестве модели предполагают, что вертикальные миграции криля на несколько сотен метров в группах глубиной в несколько десятков метров в совокупности могут создавать достаточно нисходящих струй воды, чтобы оказать значительное влияние на перемешивание океана.[66]
Плотные рои могут вызвать с жадностью есть среди хищников рыб, птиц и млекопитающих, особенно у поверхности. При потревожении стая рассеивается, а некоторые особи даже мгновенно линяют, покидая экзувия сзади как приманка.[67]
Криль обычно плавает со скоростью 5–10 см / с (2–3 длины тела в секунду),[68] использование плавательных плавательных средств для движения. Их более крупные миграции подвержены океанским течениям. Когда в опасности, они показывают реакция побега называется омар - щелкая их каудальный структуры, Тельсон и уроподы, они относительно быстро движутся назад по воде, достигая скорости в диапазоне от 10 до 27 длин тела в секунду, что для крупного криля, такого как E. superba означает около 0,8 м / с (3 фута / с).[69] Их способность плавать побудила многих исследователей классифицировать взрослого криля как микронектонический формы жизни, то есть мелкие животные, способные двигаться индивидуально против (слабых) токов. Личиночные формы криля обычно считаются зоопланктоном.[70]
Биогеохимические циклы

Антарктический криль - важный вид в контексте биогеохимический цикл[71][39] и в Антарктическая пищевая сеть.[72][73] Он играет важную роль в Южном океане из-за своей способности цикл питательных веществ и кормить пингвинов и усатый и Голубые киты.

Человеческое использование

История сбора урожая
Криль собирали как источник пищи для людей и домашних животных, по крайней мере, с 19 века, а возможно и раньше в Японии, где он был известен как окиами. Крупномасштабный промысел развился в конце 1960-х - начале 1970-х годов, и сейчас он осуществляется только в антарктических водах и в морях вокруг Японии. Исторически крупнейшими странами, ведущими промысел криля, были Япония и Советский Союз, или, после распада последнего, Россия и Украина.[74] Пик вылова, который в 1983 году составлял около 528 000 тонн только в Южном океане (из которых Советский Союз получил 93%), теперь регулируется в качестве меры предосторожности против перелова.[75]
В 1993 г. два события привели к сокращению промысла криля: Россия вышла из отрасли; и Конвенция о сохранении морских живых ресурсов Антарктики (АНТКОМ) установил максимальные квоты на вылов устойчивая эксплуатация антарктического криля. После проверки в октябре 2011 года Комиссия решила не изменять квоту.[76]
Годовой вылов в Антарктике стабилизировался на уровне около 100 000 тонн, что составляет примерно одну пятидесятую квоты на вылов АНТКОМа.[77] Основным ограничивающим фактором, вероятно, были высокие затраты наряду с политическими и правовыми проблемами.[78] Японский промысел достиг примерно 70 000 тонн.[79]
Хотя криль встречается во всем мире, промысел в Южных океанах предпочтителен, потому что криль в этих регионах более «уловлен» и его много. Особенно в антарктических морях, которые считаются первозданный, они считаются «чистым продуктом».[74]
В 2018 году было объявлено, что почти все компании, занимающиеся промыслом криля в Антарктиде, с 2020 года откажутся от операций на огромных территориях вокруг Антарктического полуострова, включая «буферные зоны» вокруг гнездящихся колоний пингвинов.[80]
Потребление человеком

Хотя общая биомасса антарктического криля может достигать 400 миллионов тонны, человеческое влияние на это краеугольные камни растет, при этом общий вылов рыбы увеличился на 39% до 294 000 тонн за 2010–2014 годы.[77] Основные страны, участвующие в промысле криля: Норвегия (56% от общего улова в 2014 г.), Республика Корея (19%), и Китай (18%).[77]
Криль - богатый источник белок и омега-3 жирные кислоты которые находятся в стадии разработки в начале 21-го века как человеческая пища, пищевые добавки в виде масляных капсул, домашний скот еда и корм для животных.[74][76][81] Криль на вкус соленый с несколько более сильным рыбным вкусом, чем у креветок. Продукты массового потребления и готовые промышленные продукты необходимо очистить от кожуры, чтобы удалить несъедобные. экзоскелет.[81]
В 2011 году США Управление по контролю за продуктами и лекарствами опубликовал письмо об отсутствии возражений в отношении изготовленного масло криля продукт быть общепризнанно безопасным (ГРАС ) для потребления человеком.[82]
Криль (и другие планктонный креветка ) наиболее широко потребляются в Юго-Восточная Азия, где это ферментированный (с неповрежденной скорлупой) и обычно мелко измельчают для получения креветочная паста. Его можно жарить и есть вместе с белым рисом или использовать для добавления умами ароматизирует широкий выбор традиционных блюд.[83][84] Жидкость процесса ферментации также собирается как рыбный соус.[85]
Смотрите также
 Портал ракообразных
Портал ракообразных- Креветки холодноводные
Рекомендации
- ^ «Криль». Интернет-словарь этимологии. Получено 22 июня, 2010.
- ^ А. Аткинсон; В. Сигель; E.A. Пахомов; М.Дж. Джессопп; В. Леб (2009). «Переоценка общей биомассы и годовой продукции антарктического криля» (PDF). Глубоководные исследования, часть I. 56 (5): 727–740. Bibcode:2009DSRI ... 56..727A. Дои:10.1016 / j.dsr.2008.12.007.
- ^ Фолькер Сигель (2011). Зигель V (ред.). "Euphausiidae Dana, 1852". Всемирная база данных Euphausiacea. Всемирный регистр морских видов. Получено 25 ноября, 2011.
- ^ Э. Бринтон (1962). «Распространение тихоокеанских эвфаузиид». Бык. Scripps Inst. Oceanogr. 8 (2): 51–270.
- ^ а б С. Николь; Ю. Эндо (1999). «Промысел криля: последствия для развития, управления и экосистемы». Водные живые ресурсы. 12 (2): 105–120. Дои:10.1016 / S0990-7440 (99) 80020-5.
- ^ а б c d Андреас Маас; Дитер Валошек (2001). "Личиночное развитие Euphausia superba Дана, 1852 г. и филогенетический анализ Euphausiacea » (PDF). Гидробиология. 448: 143–169. Дои:10.1023 / А: 1017549321961. S2CID 32997380. Архивировано из оригинал (PDF) на 2011-07-18.
- ^ Бернадетт Казанова (1984). «Филогения евфаузиак (Crustacés Eucarides)» [Филогения Euphausiacea (Crustacea: Eucarida)]. Бюллетень национального музея естественной истории (На французском). 4: 1077–1089.
- ^ а б c Бернадетт Казанова (2003). "Ordre des Euphausiacea Dana, 1852". Ракообразные. 76 (9): 1083–1121. Дои:10.1163/156854003322753439. JSTOR 20105650.
- ^ Г-н Юджиния Д'Амато; Гордон В. Харкинс; Тулио де Оливейра; Питер Р. Теске; Марк Дж. Гиббонс (2008). «Молекулярное датирование и биогеография неритического криля. Никтифан" (PDF). Морская биология. 155 (2): 243–247. Дои:10.1007 / s00227-008-1005-0. S2CID 17750015.
- ^ а б c Саймон Н. Джарман (2001). «История эволюции криля, полученная на основе анализа последовательности большой субъединицы ядер рДНК». Биологический журнал Линнеевского общества. 73 (2): 199–212. Дои:10.1111 / j.1095-8312.2001.tb01357.x.
- ^ Синь Шэнь; Хайцин Ван; Минсяо Ван; Бинь Лю (2011). "Полная последовательность митохондриального генома Евфаузия тихая (Malacostraca: Euphausiacea) обнаруживает новый порядок генов и необычные тандемные повторы ». Геном. 54 (11): 911–922. Дои:10.1139 / g11-053. PMID 22017501.
- ^ Йохан Эрик Вести Боас (1883). "Studien über die Verwandtschaftsbeziehungen der Malacostraken" [Исследования взаимоотношений Malacostraca]. Morphologisches Jahrbuch (на немецком). 8: 485–579.
- ^ а б Роберт Герни (1942). Личинки Decapod Crustacea (PDF). Луч Общество.
- ^ Изабелла Гордон (1955). «Систематическое положение Евфаузицеи». Природа. 176 (4489): 934. Bibcode:1955Натура.176..934Г. Дои:10.1038 / 176934a0. S2CID 4225121.
- ^ Триша Спирс, Рональд В. ДеБри, Лоуренс Г. Абеле и Катаржина Чодил (2005). Бойко, Кристофер Б. (ред.). «Монофилия перакарид и межпочечная филогения, выведенная из ядерных последовательностей малой субъединицы рибосомной ДНК (Crustacea: Malacostraca: Peracarida)» (PDF). Труды Биологического общества Вашингтона. 118 (1): 117–157. Дои:10.2988 / 0006-324X (2005) 118 [117: PMAIPI] 2.0.CO; 2.CS1 maint: несколько имен: список авторов (связь)
- ^ К. Меланд; Э. Уиллассен (2007). «Разобщенность« Mysidacea »(Crustacea)». Молекулярная филогенетика и эволюция. 44 (3): 1083–1104. CiteSeerX 10.1.1.653.5935. Дои:10.1016 / j.ympev.2007.02.009. PMID 17398121.
- ^ Фредерик Р. Шрам (1986). Ракообразные. Oxford University Press. ISBN 978-0-19-503742-5.
- ^ Дж. Дж. Торрес; Дж. Дж. Чайлдресс (1985). «Дыхание и химический состав батипелагической эвфаузииды. Бентуфаузия амблиопс". Морская биология. 87 (3): 267–272. Дои:10.1007 / BF00397804. S2CID 84486097.
- ^ Фолькер Сигель (2011). "Thysanoessa Брандт, 1851 г. ". Черви. Всемирный регистр морских видов. Получено 18 июня, 2011.
- ^ а б Д'Амато, М. и другие.: "Молекулярное датирование и биогеография неритического криля Никтифан ", в Морская биология об. 155, нет. 2, стр. 243-247, август 2008 г.
- ^ Фолькер Сигель (2011). В. Сигель (ред.). "Никтифан Сарс, 1883 г. ". Всемирная база данных Euphausiacea. Всемирный регистр морских видов. Получено 18 июня, 2011.
- ^ а б c Дж. Мочлайн; Л. Р. Фишер (1969). Биология эвфаузиид. Успехи в морской биологии. 7. Академическая пресса. ISBN 978-7-7708-3615-2.
- ^ а б c Хайме Гомес-Гутьеррес; Карлос Дж. Робинсон (2005). "Эмбриональное, раннее личиночное развитие, механизм вылупления и межплодный период мешочковой эвфаузииды. Никтифан симплекс Хансен ". Журнал исследований планктона. 27 (3): 279–295. Дои:10.1093 / планкт / fbi003.
- ^ С. Н. Джарман; Н. Г. Эллиотт; С. Николь; А. МакМинн (2002). "Генетическая дифференциация прибрежного антарктического криля Евфаузия кристалорфиас". Наследственность. 88 (4): 280–287. Дои:10.1038 / sj.hdy.6800041. PMID 11920136.
- ^ Р. Эскрибано; В. Марин; К. Иррибаррен (2000). "Распределение Euphausia mucronata в районе апвеллинга полуострова Меджильонес, север Чили: влияние слоя минимального содержания кислорода ». Scientia Marina. 64 (1): 69–77. Дои:10.3989 / scimar.2000.64n169.
- ^ П. Брейггеман. "Евфаузия кристалорфиас". Подводный полевой путеводитель по острову Росс и проливу Мак-Мердо, Антарктида. Калифорнийский университет в Сан-Диего.
- ^ "Криль, Euphausia superba". MarineBio.org. Получено 25 февраля, 2009.
- ^ Дж. А. Кирквуд (1984). «Путеводитель по Евфаузиаке Южного океана». Аналитические заметки ANARE. 1: 1–45.
- ^ А. Сала; М. Аззали; А. Руссо (2002). "Криль моря Росса: распространение, численность и демография Euphausia superba и Евфаузия кристалорфиас во время Итальянской антарктической экспедиции (январь – февраль 2000 г.) ». Scientia Marina. 66 (2): 123–133. Дои:10.3989 / scimar.2002.66n2123.
- ^ Г. В. Хози; М. Фукути; С. Кавагути (2003). «Разработка системы непрерывной съемки планктона в Южном океане» (PDF). Прогресс в океанографии. 58 (2–4): 263–283. Bibcode:2003PrOce..58..263H. Дои:10.1016 / j.pocean.2003.08.007.[постоянная мертвая ссылка ]
- ^ Э. Гатен. "Meganyctiphanes norvegica". Университет Лестера. Архивировано из оригинал 1 июля 2009 г.. Получено 25 февраля, 2009.
- ^ Э. Бринтон (1953). "Thysanopoda spinicauda, новый батипелагический гигантский ракообразный эвфаузиид, со сравнительными сведениями о T. Cornuta и T. egregia". Журнал Вашингтонской академии наук. 43: 408–412.
- ^ «Евфаузицея». Тасманский институт аквакультуры и рыболовства. Архивировано из оригинал 30 сентября 2009 г.. Получено 6 июня, 2010.
- ^ О. Шимомура (1995). «Роли двух крайне нестабильных компонентов F и P, участвующих в биолюминесценции эвфаузиид». Журнал биолюминесценции и хемилюминесценции. 10 (2): 91–101. Дои:10.1002 / bio.1170100205. PMID 7676855.
- ^ Дж. К. Данлэп; Дж. У. Гастингс; О. Шимомура (1980). «Перекрестная реактивность между светоизлучающими системами отдаленно родственных организмов: новый тип светоизлучающего соединения». Труды Национальной академии наук. 77 (3): 1394–1397. Bibcode:1980PNAS ... 77.1394D. Дои:10.1073 / pnas.77.3.1394. JSTOR 8463. ЧВК 348501. PMID 16592787.
- ^ П. Дж. Херринг; Э. А. Виддер (2001). «Биолюминесценция в планктоне и нектоне». В Дж. Х. Стиле; С. А. Торп; К. К. Турекян (ред.). Энциклопедия наук об океане. 1. Академическая пресса, Сан Диего. стр.308–317. ISBN 978-0-12-227430-5.
- ^ С. М. Линдси; М. И. Латц (1999). Экспериментальные доказательства люминесцентного затенения некоторыми ракообразными-эвфаузиидами. Встреча Американского общества лимнологов и океанографии (ASLO) по водным наукам. Санта-Фе.
- ^ Сёнке Йонсен (2005). «Красное и черное: биолюминесценция и окраска морских глубин» (PDF). Интегративная и сравнительная биология. 4 (2): 234–246. Дои:10.1093 / icb / 45.2.234. PMID 21676767. S2CID 247718. Архивировано из оригинал (PDF) на 2005-10-02.
- ^ а б c d Каван, Э.Л., Белчер, А., Аткинсон, А., Хилл, С.Л., Кавагути, С., МакКормак, С., Мейер, Б., Никол, С., Ратнараджа, Л., Шмидт, К. и Стейнберг, DK (2019) «Значение антарктического криля в биогеохимических циклах». Связь с природой, 10(1): 1–13. Дои:10.1038 / s41467-019-12668-7.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Г. К. Криппс; А. Аткинсон (2000). «Состав жирных кислот как показатель хищничества антарктического криля, Euphausia superba". Канадский журнал рыболовства и водных наук. 57 (S3): 31–37. Дои:10.1139 / f00-167.
- ^ а б Олав Сэтер; Тронд Эрлинг Эллингсен; Вигго Мор (1986). «Липиды североатлантического криля» (PDF). Журнал липидных исследований. 27 (3): 274–285. PMID 3734626.
- ^ М. Дж. Шрамм (10 октября 2007 г.). «Крошечный криль: гиганты в морской пищевой цепи». Национальная программа морских заповедников NOAA. Получено 4 июня, 2010.
- ^ Дж. Вейер (1999). «Меняющиеся течения окрашивают Берингово море в новый оттенок синего». NOAA Обсерватория Земли. Получено 15 июня, 2005.
- ^ Р. Д. Бродер; Г. Х. Круз; П. А. Ливингстон; Г. Уолтерс; Дж. Ианелли; Г. Л. Шварцман; М. Степаненко; Т. Уилли-Эчеверриа (1998). Проект отчета международного семинара FOCI по современным условиям в Беринговом море. NOAA. С. 22–26.
- ^ Дж. Роуч (17 июля 2003 г.). «Ученые обнаружили загадочного убийцу криля». National Geographic News.
- ^ Х. Гомес-Гутьеррес; В. Т. Петерсон; А. де Роберти; Р. Д. Бродер (2003). «Массовая гибель криля от паразитоидных инфузорий». Наука. 301 (5631): 339. Дои:10.1126 / science.1085164. PMID 12869754. S2CID 28471713.
- ^ Дж. Д. Шилдс; Х. Гомес-Гутьеррес (1996). "Oculophryxus bicaulis, новый род и вид равноногих даджидов, паразитирующих на эвфаузииде. Стилохейрон аффинный Хансен ». Международный журнал паразитологии. 26 (3): 261–268. Дои:10.1016/0020-7519(95)00126-3. PMID 8786215.
- ^ Расти Дорнин (6 июля 1997 г.). «Популяции антарктического криля сокращаются». CNN. Получено 18 июня, 2011.
- ^ Доусон, Аманда Л; Кавагути, Со; Король, Екатерина К; Таунсенд, Кэти А; Кинг, Роберт; Huston, Wilhelmina M; Бенгтсон Нэш, Сьюзан М (2018). «Превращение микропластика в нанопластик путем фрагментации пищеварительной системы антарктическим крилем». Nature Communications. 9 (1): 1001. Bibcode:2018НатКо ... 9.1001D. Дои:10.1038 / s41467-018-03465-9. ЧВК 5843626. PMID 29520086.
- ^ М. Д. Найт (1984). "Изменения в морфогенезе личинок в популяции южной Калифорнийской бухты Евфаузия тихая с зимы по лето 1977–1978 " (PDF). Отчет CalCOFI. XXV.
- ^ "Euphausia superba". Информационный бюллетень по видам. Продовольственная и сельскохозяйственная организация. Получено 4 июня, 2010.
- ^ Р. М. Росс; Л. Б. Кветин (1986). «Насколько продуктивен антарктический криль?». Бионаука. 36 (4): 264–269. Дои:10.2307/1310217. JSTOR 1310217.
- ^ Жанин Кузен-Руди (2000). "Сезонное воспроизводство, многократный нерест и плодовитость северного криля, Meganyctiphanes norvegicaи антарктический криль, Euphausia superba". Канадский журнал рыболовства и водных наук. 57 (S3): 6–15. Дои:10.1139 / f00-165.
- ^ Х. Гомес-Гутьеррес (2002). «Механизм вылупления и отложенное вылупление яиц трех нерестовых видов эвфаузиид в лабораторных условиях». Журнал исследований планктона. 24 (12): 1265–1276. Дои:10.1093 / планкт / 24.12.1265.
- ^ Э. Бринтон; М. Д. Оман; А. В. Таунсенд; М. Д. Найт; А. Л. Бриджмен (2000). Евфаузииды Мирового океана. Серия компакт-дисков Всемирной базы данных по биоразнообразию, Springer Verlag. ISBN 978-3-540-14673-5.
- ^ Ф. Бухгольц (2003). «Эксперименты по физиологии южного и северного криля, Euphausia superba и Meganyctiphanes norvegica, с акцентом на линьку и рост - обзор ». Поведение и физиология в морской и пресноводной среде. 36 (4): 229–247. Дои:10.1080/10236240310001623376. S2CID 85121989.
- ^ Х.-К. Голень; С. Николь (2002). "Использование зависимости между диаметром глаза и длиной тела для выявления последствий длительного голодания для антарктического криля. Euphausia superba". Серия "Прогресс морской экологии". 239: 157–167. Bibcode:2002MEPS..239..157S. Дои:10,3354 / meps239157.
- ^ Б. Маринович; М. Мангель (1999). «Криль может уменьшиться в результате экологической адаптации к временно неблагоприятной окружающей среде» (PDF). Письма об экологии. 2: 338–343.
- ^ Х. Г. Гомес (1995). "Распространение, численность и динамика численности эвфаузиид.Никтифан симплекс и Евфаузия эксимия у западного побережья Нижней Калифорнии, Мексика " (PDF). Серия "Прогресс морской экологии". 119: 63–76. Bibcode:1995MEPS..119 ... 63G. Дои:10.3354 / meps119063.
- ^ У. Килс; П. Маршалл (1995). "Der Krill, wie er schwimmt und frisst - neue Einsichten mit neuen Methoden ("Антарктический криль - как он плавает и питается - новые открытия с помощью новых методов")". У И. Хемпеля; Г. Хемпель (ред.). Biologie der Polarmeere - Erlebnisse und Ergebnisse (Биология полярных океанов: опыт и результаты). Фишер Верлаг. С. 201–210. ISBN 978-3-334-60950-7.
- ^ Р. Пайпер (2007). Необычные животные: энциклопедия любопытных и необычных животных. Гринвуд Пресс. ISBN 978-0-313-33922-6.
- ^ Gandomi, A.H .; Алави, А.Х. (2012). «Стадо криля: новый алгоритм оптимизации, вдохновленный биологией». Коммуникации в нелинейной науке и численном моделировании. 17 (12): 4831–4845. Bibcode:2012CNSNS..17.4831G. Дои:10.1016 / j.cnsns.2012.05.010.
- ^ Дж. С. Джаффе; М. Д. Оманн; А. де Роберти (1999). "Сонарные оценки уровней дневной активности Евфаузия тихая в бухте Саанич » (PDF). Канадский журнал рыболовства и водных наук. 56 (11): 2000–2010. Дои:10.1139 / cjfas-56-11-2000. Архивировано из оригинал (PDF) на 2011-07-20.
- ^ Герайнт А. Тарлинг; Магнус Л. Джонсон (2006). «Сытость вызывает у криля чувство опускания». Текущая биология. 16 (3): 83–84. Дои:10.1016 / j.cub.2006.01.044. PMID 16461267.
- ^ Дэн Ховард (2001). «Криль» (PDF). У Германа А. Карла; Джон Л. Чин; Эдвард Юбер; Питер Х. Штауфер; Джеймс В. Хендли II (ред.). За Золотыми воротами - океанография, геология, биология и экологические проблемы в заливе Фараллонес. Геологическая служба США. С. 133–140. Циркуляр 1198. Получено 8 октября, 2011.
- ^ Уишарт, Скай (июль – август 2018 г.). «Эффект криля». New Zealand Geographic (152): 24.
- ^ Д. Ховард. «Криль в национальном морском заповеднике Корделл-Бэнк». Национальное управление океанических и атмосферных исследований. Получено 15 июня, 2005.
- ^ Дэвид А. Демер; Стефан Г. Конти (2005). «Новая модель силы цели указывает на большее количество криля в Южном океане». Журнал морских наук ICES. 62 (1): 25–32. Дои:10.1016 / j.icesjms.2004.07.027.
- ^ У. Килс (1982). «Плавательное поведение, плавательные способности и энергетический баланс антарктического криля Euphausia superba". Серия исследований БИОМАССЫ 3, Серия исследований БИОМАСС: 1–122.
- ^ С. Николь; Ю. Эндо (1997). "Мировые промыслы криля". Технический документ ФАО по рыболовству. 367.
- ^ Ратнараджа, Л., Боуи, А.Р., Ланнузель, Д., Майнерс, К. и Никол С. (2014) «Биогеохимическая роль усатых китов и криля в круговороте питательных веществ Южного океана». PLOS ONE, 9(12): e114067. Дои:10.1371 / journal.pone.0114067
- ^ Хопкинс, Т.Л., Эйнли, Д.Г., Торрес, Дж. Дж., Ланкрафт, Т.М., 1993. Трофическая структура в открытых водах краевой ледовой зоны в районе слияния Скотия-Уэдделл весной (1983 г.) Полярная биология 13, 389–397.
- ^ Lancraft, T.M., Relsenbichler, K.R., Robinson, B.H., Hopkins, T.L., Torres, J.J., 2004. Сообщество микронектона и макрозоопланктона с преобладанием криля в проливе Крокер, Антарктида, с оценкой хищничества рыб. Deep-Sea Research II 51, 2247–2260.
- ^ а б c Гроссман, Элизабет (14 июля 2015 г.). «Ученые рассматривают необходимость защиты криля от чрезмерной охоты человека». Международное общественное радио (PRI). Получено 1 апреля 2017.
- ^ «Промысел криля и устойчивость: антарктический криль (Euphausia superba)». Комиссия по сохранению морских живых ресурсов Антарктики. 23 апреля 2015 г.. Получено 1 апреля 2017.
- ^ а б Ширмайер, Q (2010). «Экологи опасаются кризиса с антарктическим крилем». Природа. 467 (7311): 15. Дои:10.1038 / 467015a. PMID 20811427.
- ^ а б c «Криль - биология, экология и рыболовство». Комиссия по сохранению морских живых ресурсов Антарктики. 28 апреля 2015 г.. Получено 1 апреля 2017.
- ^ Минтурн Дж. Райт (1987). «Право собственности на Антарктиду, ее живые и минеральные ресурсы». Журнал права и окружающей среды. 4 (2): 49–78.
- ^ С. Николь; Дж. Фостер (2003). «Последние тенденции в промысле антарктического криля». Водные живые ресурсы. 16: 42–45. Дои:10.1016 / S0990-7440 (03) 00004-4.
- ^ Джош, Габбатисс (10 июля 2018 г.). «Индустрия криля поддерживает огромный заповедник в Антарктическом океане для защиты пингвинов, тюленей и китов». Независимый. Получено 10 июля 2018.
- ^ а б "Почему криль?". Юго-западный научный центр рыболовства, Национальное управление океанических и атмосферных исследований США. 22 ноября 2016 г.. Получено 1 апреля 2017.
- ^ Чизмен М.А. (22 июля 2011 г.). «Масло криля: Ответное письмо Агентства, уведомление GRAS № GRN 000371». США FDA. Получено 3 июн 2015.
- ^ Омори, М. (1978). «Мировой зоопланктонный промысел: обзор». Морская биология. 48 (3): 199–205. Дои:10.1007 / BF00397145. S2CID 86540101.
- ^ Понгсеткул, Джаксума; Бенджакул, Соттават; Сампавапол, Пуннани; Осако, Кадзуфуми; Фэйтонг, Нандша (17 сентября 2014 г.). «Химический состав и физические свойства соленой креветочной пасты (Капи) производства Таиланда». Международные водные исследования. 6 (3): 155–166. Дои:10.1007 / s40071-014-0076-4.
- ^ Абэ, Кенджи; Судзуки, Кенджи; Хашимото, Канехиса (1979). «Использование криля в качестве материала для рыбного соуса». Ниппон Суисан Гаккаиси. 45 (8): 1013–1017. Дои:10.2331 / suisan.45.1013.
дальнейшее чтение
- Боден, Брайан П .; Джонсон, Мартин В.; Бринтон, Эдвард: «Euphausiacea (Crustacea) северной части Тихого океана». Бюллетень Океанографического института Скриппса. Том 6, номер 8, 1955 год.
- Бринтон, Эдвард: «Евфаузииды вод Юго-Восточной Азии». Отчет Нага том 4, часть 5. Ла-Хойя: Калифорнийский университет, Океанографический институт Скриппса, 1975 г.
- Конвей, Д. В. П .; White, R.G .; Hugues-Dit-Ciles, J .; Galienne, C.P .; Робинс, Д. Б .: Путеводитель по прибрежному и поверхностному зоопланктону юго-западной части Индийского океана, Заказ Euphausiacea, Периодическая публикация Морская биологическая ассоциация Соединенного Королевства № 15, Плимут, Великобритания, 2003 г.
- Эверсон, И. (ред.): Криль: биология, экология и рыболовство. Оксфорд, Blackwell Science; 2000 г. ISBN 0-632-05565-0.
- Хамнер, Уильям М. (май 1984 г.). «Криль - неиспользованная добыча с моря?». Национальная география. Vol. 165 нет. 5. С. 626–642. ISSN 0027-9358. OCLC 643483454.
- Mauchline, J .: Euphausiacea: взрослые люди, Международный совет по разведке моря, 1971. Идентификационные листы взрослой особи криля с множеством штриховых рисунков. PDF напильник, 2МБ.
- Mauchline, J .: Euphausiacea: Личинки, Conseil International pour l'Exploration de la Mer, 1971. Идентификационные листы личиночных стадий криля с множеством штриховых рисунков. PDF файл, 3 Мб.
- Тетт, П .: Биология эвфаузиид, конспекты лекций 2003 курс по морской биологии из Университет Напьера.
- Тетт, П .: Биолюминесценция, конспекты лекций из выпуска 1999/2000 того же курса.
внешняя ссылка
- Веб-камера Крилевого аквариума в Австралийском антарктическом отделе
- "Энергия Антарктики" анимация Лизы Робертс

Основной коммерческий рыболовство видовые группы | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дикий |
|    | ||||||||||||||||
| На ферме | ||||||||||||||||||