Экосистема озера - Lake ecosystem
А экосистема озера или озерная экосистема включает в себя биотический (живущий) растения, животные и микроорганизмы, а также абиотический (неживые) физические и химические взаимодействия.[1] Экосистемы озер являются ярким примером непроточные экосистемы (лентичный относится к стационарным или относительно неподвижным пресная вода, от латинский лентус, что означает "вялый"), в том числе пруды, озера и водно-болотные угодья, и большая часть этой статьи относится к лентичным экосистемам в целом. Бледные экосистемы можно сравнить с лотические экосистемы, которые связаны с проточными земными водами, такими как реки и потоки. Вместе эти две области образуют более общую область изучения пресная вода или водная экология.
Лентичные системы разнообразны: от небольшого временного бассейна с дождевой водой глубиной в несколько дюймов до озеро Байкал, максимальная глубина которого составляет 1642 м.[2] Общее различие между бассейнами / прудами и озерами нечеткое, но Браун[1] утверждает, что вся нижняя поверхность прудов и бассейнов освещена светом, а озера - нет. Кроме того, некоторые озера становятся сезонно стратифицированными (более подробно это обсуждается ниже). Пруды и бассейны имеют две области: пелагический зона открытой воды, а бентосная зона, включающий в себя нижнюю и прибрежную области. Поскольку у озер есть глубокие участки дна, не освещенные светом, в этих системах есть дополнительная зона, глубокий.[3] Эти три области могут иметь очень разные абиотические условия и, следовательно, виды-хозяева, которые специально приспособлены к жизни там.[1]
Важные абиотические факторы
Свет
Свет обеспечивает солнечную энергию, необходимую для управления процессом фотосинтез, главный источник энергии лентичных систем.[2] Количество получаемого света зависит от комбинации нескольких факторов. Небольшие пруды могут затеняться окружающими деревьями, а облачный покров может влиять на доступность света во всех системах, независимо от размера. Сезонные и суточные условия также влияют на доступность света, потому что чем меньше угол, под которым свет падает на воду, тем больше света теряется при отражении. Это известно как Закон пива.[4] Как только свет проникает через поверхность, он может также рассеиваться частицами, взвешенными в толще воды. Это рассеяние уменьшает общее количество света по мере увеличения глубины.[3][5] Озера делятся на фотический и афотический регионы, в которых ранее получал солнечный свет, а последний находится ниже глубины проникновения света, что лишает их фотосинтетической способности.[2] Что касается зонирования озер, считается, что пелагическая и бентическая зоны находятся в пределах фотической области, а профундальная зона - в афотической.[1]
Температура
Температура является важным абиотическим фактором в непроточных экосистемах, поскольку большая часть биоты пойкилотермный, где внутренняя температура тела определяется окружающей системой. Вода может быть нагрета или охлаждена за счет излучения на поверхности и проводимости к или от воздуха и окружающей подложки.[4] Мелкие водоемы часто имеют постоянный температурный градиент от более теплой воды на поверхности до более холодной воды на дне. Кроме того, колебания температуры в этих системах могут сильно различаться как в дневное, так и в сезонное время.[1]
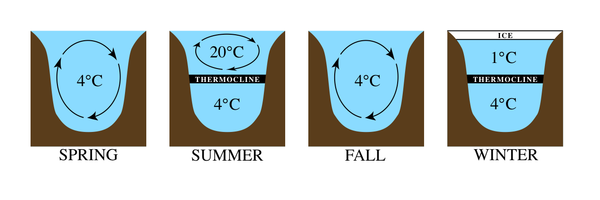
Температурные режимы больших озер очень разные (рис. 2). Например, в регионах с умеренным климатом при повышении температуры воздуха ледяной слой, образовавшийся на поверхности озера, распадается, оставляя воду с температурой около 4 ° C. Это температура, при которой вода имеет самую высокую плотность. В течение сезона более теплые температуры воздуха нагревают поверхностные воды, делая их менее плотными. Более глубокие воды остаются прохладными и плотными из-за меньшего проникновения света. С началом лета образуются два отдельных слоя с такой большой разницей температур между ними, что они остаются стратифицированными. Самая низкая зона в озере самая холодная и называется гиполимнион. Верхняя теплая зона называется эпилимнион. Между этими зонами простирается полоса быстрого изменения температуры, называемая термоклин. В более холодное осеннее время тепло теряется на поверхности, и эпилимнион охлаждается. Когда температуры в двух зонах становятся достаточно близкими, воды снова начинают перемешиваться, чтобы создать однородную температуру, и это событие называется водоворот озера. Зимой происходит обратная стратификация, так как вода у поверхности остывает замерзает, а более теплая, но более плотная вода остается у дна. Устанавливается термоклин, цикл повторяется.[1][2]
ветер

В открытых системах ветер может создавать турбулентные, спиралевидные поверхностные токи, называемые Тиражи Ленгмюра (Рис. 3). Как именно эти токи возникают, до сих пор не совсем понятно, но очевидно, что это связано с некоторым взаимодействием между горизонтальными поверхностными токами и поверхностными гравитационными волнами. Видимый результат этих вращений, который можно увидеть в любом озере, - линии пены на поверхности, идущие параллельно направлению ветра. Положительно плавучие частицы и мелкие организмы концентрируются в линии пены у поверхности, а отрицательно плавучие объекты обнаруживаются в восходящем течении между двумя вращениями. Объекты с нейтральной плавучестью обычно равномерно распределяются в толще воды.[2][3] Эта турбулентность обеспечивает циркуляцию питательных веществ в водной толще, что делает ее критически важной для многих пелагических видов, однако ее влияние на бентические и глубинные организмы соответственно минимально или отсутствует.[3] Степень циркуляции питательных веществ зависит от системы, поскольку она зависит от таких факторов, как сила и продолжительность ветра, а также глубина озера или бассейна и продуктивность.
Химия
Кислород необходим для организма дыхание. Количество кислорода в стоячей воде зависит от: 1) площади прозрачной воды, подверженной воздействию воздуха, 2) циркуляции воды в системе и 3) количества кислорода, производимого и используемого присутствующими организмами.[1] В неглубоких, богатых растениями водоемах могут быть большие колебания содержания кислорода, с чрезвычайно высокими концентрациями в течение дня из-за фотосинтеза и очень низкими значениями ночью, когда дыхание является доминирующим процессом первичных продуцентов. Термическая стратификация в более крупных системах также может влиять на количество кислорода, присутствующего в различных зонах. Эпилимнион богат кислородом, потому что он быстро циркулирует, получая кислород через контакт с воздухом. Гиполимнион, однако, циркулирует очень медленно и не контактирует с атмосферой. Кроме того, в гиполимнионе меньше зеленых растений, поэтому в результате фотосинтеза выделяется меньше кислорода. Весной и осенью, когда эпилимнион и гиполимнион смешиваются, кислород становится более равномерно распределенным в системе. Низкий уровень кислорода характерен для профундальной зоны из-за скопления разлагающейся растительности и животного материала, который «проливается дождем» из пелагических и бентосных зон, и неспособности поддерживать первичных продуцентов.[1]
Фосфор важен для всех организмов, потому что он является компонентом ДНК и РНК и участвует в метаболизме клетки как компонент АТФ и АДФ. Кроме того, фосфор не содержится в больших количествах в пресноводных системах, что ограничивает фотосинтез в первичных продуцентах, что делает его основным фактором, определяющим производство постноводных систем. Цикл фосфора сложен, но модель, описанная ниже, описывает основные пути. Фосфор в основном попадает в пруд или озеро через сток с водосбора или атмосферные осадки. Попадая в систему, реактивная форма фосфора обычно поглощается водорослями и макрофитами, которые выделяют нереактивное соединение фосфора в качестве побочного продукта фотосинтеза. Этот фосфор может дрейфовать вниз и стать частью донных или глубоких отложений, или он может быть реминерализованный в реактивную форму микробами в толще воды. Точно так же инертный фосфор в осадке может быть реминерализован в реактивную форму.[2] Однако отложения обычно богаче фосфором, чем вода в озере, что указывает на то, что это питательное вещество может долго оставаться там, прежде чем оно будет реминерализовано и повторно введено в систему.[3]
Лентиочная системная биота
Бактерии
Бактерии присутствуют во всех районах непроточных вод. Свободноживущие формы связаны с разложением органического материала, биопленка на поверхности скал и растений, взвешенных в толще воды, а также в отложениях придонной и профундальной зон. Другие формы также связаны с кишечником непереносимых животных как паразиты или комменсальный отношения.[3] Бактерии играют важную роль в системном метаболизме за счет рециркуляции питательных веществ,[2] который обсуждается в разделе «Трофические отношения».
Первичные производители

Водоросли, включая оба фитопланктон и перифитон, являются основными фотосинтезаторами в прудах и озерах. Фитопланктон дрейфует в водной толще пелагиали. Многие виды имеют более высокую плотность, чем вода, что должно привести к их непреднамеренному погружению в бентос. Чтобы бороться с этим, фитопланктон разработал механизмы изменения плотности, образуя вакуоли и газовые пузырьки, или изменяя их форму, чтобы вызвать сопротивление, замедляя тем самым их спуск. Очень сложная адаптация, используемая небольшим количеством видов, - это хвостовидный жгутик который может регулировать вертикальное положение и позволять движение в любом направлении.[2] Фитопланктон также может поддерживать свое присутствие в водной толще, циркулируя в Ленгмюровские вращения.[3] А перифитные водоросли прикреплены к субстрату. В озерах и прудах они могут покрывать все поверхности бентоса. Оба типа планктона важны как источники пищи и как поставщики кислорода.[2]
Водные растения живут как в придонной, так и в пелагической зонах и могут быть сгруппированы по способу роста: ⑴ возникающий = укореняется в субстрате, но листья и цветы уходят в воздух; ⑵ с плавающими листьями = укореняется в субстрате, но с плавающими листьями; ⑶ погруженный = растет под поверхностью; ⑷ свободно плавающие макрофиты = не укореняется в субстрате, а плавает на поверхности.[1] Эти различные формы макрофитов обычно встречаются в разных частях бентосной зоны, с зарождающейся растительностью ближе всего к береговой линии, затем с плавающими листьями макрофитов, а затем с подводной растительностью. Свободно плавающие макрофиты могут встречаться где угодно на поверхности системы.[2]
Водные растения обладают большей плавучестью, чем их наземные аналоги, потому что пресная вода имеет более высокую плотность, чем воздух. Это делает конструктивную жесткость не важной в озерах и прудах (за исключением воздушных стеблей и листьев). Таким образом, листья и стебли большинства водных растений используют меньше энергии для создания и поддержания древесной ткани, вместо этого вкладывая эту энергию в быстрый рост.[1] Чтобы противостоять стрессам, вызываемым ветром и волнами, растения должны быть гибкими и жесткими. Свет, глубина воды и типы субстрата являются наиболее важными факторами, контролирующими распространение подводных водных растений.[6] Макрофиты являются источниками пищи, кислорода и структуры среды обитания в бентосной зоне, но не могут проникать в глубины эвфотической зоны и, следовательно, там не встречаются.[1][5]
Беспозвоночные

Зоопланктон это крошечные животные, подвешенные в толще воды. Подобно фитопланктону, эти виды разработали механизмы, которые не позволяют им погружаться в более глубокие воды, включая формы тела, вызывающие сопротивление, и активное движение придатков (например, антенн или шипов).[1] Пребывание в толще воды может иметь свои преимущества с точки зрения питания, но отсутствие рефугиумов в этой зоне делает зоопланктон уязвимым для хищников. В ответ некоторые виды, особенно Дафния sp., совершают ежедневные вертикальные миграции в толще воды, пассивно опускаясь на более темные более низкие глубины днем и активно продвигаясь к поверхности ночью. Кроме того, поскольку условия в линзовой системе могут сильно меняться в зависимости от сезона, зоопланктон может переключаться с откладки обычных яиц на покоящиеся яйца, когда не хватает пищи, температура опускается ниже 2 ° C или если численность хищников высока. Эти покоящиеся яйца имеют диапауза, или период покоя, который должен позволить зоопланктону столкнуться с условиями, более благоприятными для выживания, когда они, наконец, вылупятся.[7] Среди беспозвоночных, населяющих бентическую зону, преобладают мелкие виды, и они богаты видами по сравнению с зоопланктоном открытой воды. Они включают: Ракообразные (например. крабы, рак, и креветка ), моллюски (например. моллюски и улитки ) и многочисленные виды насекомых.[2] Эти организмы в основном встречаются в зонах роста макрофитов, где находятся самые богатые ресурсы, вода с высоким содержанием кислорода и самая теплая часть экосистемы. Структурно разнообразные слои макрофитов являются важными участками накопления органического вещества и представляют собой идеальную область для колонизации. Отложения и растения также обеспечивают хорошую защиту от хищных рыб.[3]
Очень немногие беспозвоночные могут жить в холоде, темноте и бедности кислородом. профундальная зона. Те, что могут, часто имеют красный цвет из-за наличия большого количества гемоглобин, что значительно увеличивает количество кислорода, переносимого в клетки.[1] Поскольку концентрация кислорода в этой зоне низкая, большинство видов строят туннели или норы, в которых они могут спрятаться, и используют минимальное количество движений, необходимых для циркуляции воды, притягивая к себе кислород, не затрачивая слишком много энергии.[1]
Рыба и другие позвоночные
У рыб есть ряд физиологических толерантностей, которые зависят от того, к какому виду они принадлежат. У них разные смертельные температуры, потребности в растворенном кислороде и потребности нереста, которые зависят от их уровня активности и поведения. Поскольку рыбы очень подвижны, они могут бороться с неподходящими абиотическими факторами в одной зоне, просто перемещаясь в другую. Питатель детрита в профундальной зоне, например, обнаружив, что концентрация кислорода упала слишком низко, может подавать ближе к бентосной зоне. Рыба также может менять свое место жительства на разных этапах своей жизненной истории: вылупляясь из осадочного гнезда, затем перемещаясь в заросшую водорослями бентическую зону, чтобы развиваться в защищенной среде с пищевыми ресурсами, и, наконец, во взрослом возрасте в пелагиали.
Другие таксоны позвоночных также населяют линтические системы. Они включают амфибии (например. саламандры и лягушки ), рептилии (например. змеи, черепахи, и аллигаторы ), и большое количество водоплавающая птица виды.[5] Большинство этих позвоночных проводят часть своего времени в наземной среде обитания и, таким образом, не подвергаются прямому воздействию абиотических факторов в озере или пруду. Многие виды рыб важны и как потребители, и как виды добычи для более крупных позвоночных, упомянутых выше.
Трофические отношения
Первичные производители
Бледные системы получают большую часть своей энергии от фотосинтеза, выполняемого водными растениями и водорослями. Эта автохтонный Процесс включает комбинацию углекислого газа, воды и солнечной энергии для производства углеводов и растворенного кислорода. В озере или пруду потенциальная скорость фотосинтеза обычно уменьшается с глубиной из-за ослабления света. Фотосинтез, однако, часто бывает слабым на верхних нескольких миллиметрах поверхности, вероятно, из-за торможения ультрафиолетовым светом. Точные измерения глубины и скорости фотосинтеза на этой кривой зависят от системы и зависят от: 1) общей биомассы фотосинтезирующих клеток, 2) количества светопоглощающих материалов и 3) количества и частотного диапазона светопоглощающих пигментов (т.е. хлорофиллы ) внутри фотосинтезирующих клеток.[5] Энергия, создаваемая этими первичными производителями, важна для общества, потому что она передается в высшие трофические уровни через потребление.
Бактерии
Подавляющее большинство бактерий в озерах и прудах получают энергию за счет разложения растительности и животных. В пелагиали мертвая рыба и иногда аллохтонный поступления опадной подстилки являются примерами крупных твердых частиц органического вещества (CPOM> 1 мм). Бактерии разлагают их на мелкие частицы органического вещества (FPOM <1 мм), а затем на полезные питательные вещества. Мелкие организмы, такие как планктон, также характеризуются как FPOM. При разложении выделяются очень низкие концентрации питательных веществ, поскольку бактерии используют их для создания собственной биомассы. Однако бактерии потребляются простейшие, которые, в свою очередь, потребляются зоопланктоном, а затем трофические уровни. Питательные вещества, включая те, которые содержат углерод и фосфор, повторно попадают в толщу воды в любом количестве точек этой пищевой цепи через экскрецию или смерть организма, делая их снова доступными для бактерий. Этот цикл регенерации известен как микробная петля и является ключевым компонентом непроточных пищевых сетей.[2]
Разложение органических материалов может продолжаться в бентосной и глубинной зонах, если вещество проходит через толщу воды до того, как полностью переваривается пелагическими бактериями. Больше всего бактерий здесь в отложениях, где они обычно в 2-1000 раз больше, чем в толще воды.[7]
Бентические беспозвоночные
Бентические беспозвоночные из-за высокого уровня видового богатства имеют множество способов поимки добычи. Питатели фильтра создают токи через сифоны или биение ресничек, чтобы тянуть воду и ее питательные вещества к себе для напряжения. Grazers используйте приспособления для соскабливания, расчесывания и измельчения для питания перифитных водорослей и макрофитов. Члены гильдии коллекционеров просматривают отложения, выбирая конкретные частицы с хищными придатками. Кормление депозита беспозвоночные без разбора потребляют отложения, переваривая любые содержащиеся в них органические вещества. Наконец, некоторые беспозвоночные относятся к хищник гильдия, отлов и поедание живых животных.[2][8] Глубокая зона является домом для уникальной группы фильтрующих питателей, которые используют небольшие движения тела, чтобы провести ток через норы, созданные ими в отложениях. Этот режим питания требует наименьшего количества движений, позволяя этим видам экономить энергию.[1] Небольшое количество таксонов беспозвоночных являются хищниками в профундальной зоне. Эти виды, вероятно, из других регионов и заходят на эти глубины только для кормления. Подавляющее большинство беспозвоночных в этой зоне питаются отложениями, получая энергию из окружающих отложений.[8]
Рыбы
Размер, подвижность и сенсорные способности рыб позволяют им использовать широкую базу добычи, охватывающую несколько зон зональности. Как и у беспозвоночных, рыбные привычки кормления можно разделить на группы. В пелагиали травоядные животные пасутся на перифитоне и макрофитах или собирают фитопланктон из водной толщи. Хищники включают рыб, питающихся зоопланктоном в толще воды (зоопланкоядные ), насекомых на поверхности воды, на донных структурах или в донных отложениях (насекомоядные ), и те, которые питаются другой рыбой (рыбоядные ). Рыбы, которые потребляют детрит и получают энергию за счет переработки его органического материала, называются детритофаги. Всеядные животные поедают разнообразную добычу, включая растительный, фаунистический и обломочный материал. Наконец, члены паразитический Гильдия получает питание от вида-хозяина, обычно от другой рыбы или крупного позвоночного животного.[2] Таксоны рыб гибки в своих кормовых ролях, меняя свой рацион в зависимости от условий окружающей среды и наличия добычи. Многие виды также претерпевают изменения в диете по мере развития. Следовательно, вполне вероятно, что любая рыба занимает несколько гильдий кормления в течение своей жизни.[9]
Лентиочные пищевые сети
Как отмечалось в предыдущих разделах, непроточная биота связана сложной сетью трофических отношений. Можно считать, что эти организмы слабо связаны с конкретными трофическими группами (например, первичные продуценты, травоядные, первичные плотоядные, вторичные плотоядные и т. Д.). Ученые разработали несколько теорий, чтобы понять механизмы, контролирующие численность и разнообразие в этих группах. В общем, низходящий процессы диктуют, что изобилие таксонов добычи зависит от действий потребителей из более высоких трофические уровни. Обычно эти процессы протекают только между двумя трофическими уровнями, не влияя на другие. Однако в некоторых случаях водные системы испытывают трофический каскад; например, это может произойти, если первичные производители меньше страдают от выпаса травоядных животных, потому что эти травоядные животные подавляются плотоядными. Вверх дном процессы функционируют, когда изобилие или разнообразие членов более высоких трофических уровней зависит от наличия или качества ресурсов с более низких уровней. Наконец, комбинированная теория регулирования, снизу вверх: сверху вниз, сочетает в себе прогнозируемое влияние потребителей и доступность ресурсов. Он предсказывает, что трофические уровни, близкие к самым низким трофическим уровням, будут больше всего подвержены влиянию восходящих сил, в то время как нисходящие эффекты должны быть наиболее сильными на верхних уровнях.[2]
Модели сообщества и разнообразие
| Озерные зоны |
|---|
| Стратификация озера |
| Типы озер |
| Смотрите также |
Богатство местных видов
Биоразнообразие непроточной системы увеличивается с увеличением площади озера или пруда. Это связано с более высокой вероятностью обнаружения частично наземными видами более крупной системы. Кроме того, поскольку более крупные системы обычно имеют большую популяцию, вероятность исчезновения снижается.[10] Дополнительные факторы, включая температурный режим, pH, доступность питательных веществ, сложность среды обитания, скорость видообразования, конкуренцию и хищничество, были связаны с количеством видов, присутствующих в системах.[2][6]
Модели сукцессии в планктонных сообществах - модель PEG
Сообщества фитопланктона и зоопланктона в озерных системах подвергаются сезонной смене в зависимости от доступности питательных веществ, хищничества и конкуренции. Sommer и другие.[11] описал эти закономерности как часть группы Plankton Ecology Group (ПЭГ ) модель с 24 утверждениями, построенными на основе анализа многочисленных систем. Следующее включает подмножество этих утверждений, как объяснили Брёнмарк и Ханссон.[2] иллюстрируя преемственность через один сезонный цикл:
Зима
1. Повышенная доступность питательных веществ и света приводит к быстрому росту фитопланктона к концу зимы. Преобладающие виды, такие как диатомовые водоросли, имеют небольшие размеры и обладают способностью к быстрому росту. 2. Этот планктон потребляется зоопланктоном, который становится доминирующим таксоном планктона.
весна
3. А фаза чистой воды происходит по мере того, как популяции фитопланктона истощаются из-за увеличения хищничества со стороны растущего числа зоопланктона.
Лето
4. Численность зоопланктона снижается в результате сокращения добычи фитопланктона и увеличения хищничества молоди рыб.
5. С увеличением доступности питательных веществ и уменьшением хищничества со стороны зоопланктона развивается разнообразное сообщество фитопланктона.
6. По мере того, как лето продолжается, питательные вещества истощаются в предсказуемом порядке: фосфор, кремнезем, а потом азот. Численность различных видов фитопланктона варьируется в зависимости от их биологической потребности в этих питательных веществах.
7. Мелкий зоопланктон становится доминирующим типом зоопланктона, потому что он менее уязвим для хищничества рыб.
Осень
8. Хищничество рыб уменьшается из-за более низких температур и увеличения численности зоопланктона всех размеров.
Зима
9. Низкие температуры и ограниченная доступность света приводят к снижению темпов первичной продукции и сокращению популяций фитопланктона. Воспроизводство зоопланктона снижается из-за более низких температур и меньшего количества добычи.
Модель PEG представляет собой идеализированную версию этой модели сукцессии, в то время как естественные системы известны своей вариативностью.[2]
Широтные узоры
Существует хорошо задокументированная глобальная закономерность, которая коррелирует уменьшение разнообразия растений и животных с увеличением широты, то есть количество видов уменьшается по мере продвижения к полюсам. Причина этой закономерности - одна из самых больших загадок для экологов сегодня. Теории для его объяснения включают доступность энергии, климатическую изменчивость, беспокойство, конкуренцию и т. Д.[2] Несмотря на этот глобальный градиент разнообразия, эта модель может быть слабой для пресноводных систем по сравнению с глобальными морскими и наземными системами.[12] Это может быть связано с размером, поскольку Гиллебранд и Азовский[13] обнаружили, что более мелкие организмы (простейшие и планктон) не сильно следовали ожидаемой тенденции, в то время как более крупные виды (позвоночные) следовали. Они объяснили это лучшей способностью к распространению более мелкими организмами, что может привести к высокому распространению во всем мире.[2]
Жизненные циклы природных озер
Создание озера
Озера могут быть образованы разными способами, но наиболее распространенные из них кратко описаны ниже. Самые старые и крупнейшие системы являются результатом тектонический деятельность. Например, рифтовые озера в Африке являются результатом сейсмической активности на участке разделения двух тектонических плит. Ледяные озера образуются, когда ледники отступают, оставляя за собой аномалии формы ландшафта, которые затем заполняются водой. В заключение, старицы находятся речной по происхождению, в результате чего извилистая излучина реки отделяется от основного русла.[2]
Естественное вымирание
Все озера и пруды получают наносы. Поскольку эти системы на самом деле не расширяются, логично предположить, что они будут становиться все более мелкими по глубине, в конечном итоге превратившись в водно-болотные угодья или наземную растительность. Продолжительность этого процесса должна зависеть от комбинации глубины и скорости осаждения. Мох[5] приводит пример Озеро Танганьика, который достигает глубины 1500 м и имеет скорость осаждения 0,5 мм / год. Если предположить, что на осадконакопление не влияют антропогенные факторы, эта система должна исчезнуть примерно через 3 миллиона лет. Неглубокие чечевичные системы также могут заполняться, когда болота вторгаются внутрь с краев. Эти процессы протекают в гораздо более короткие сроки, требуя от сотен до тысяч лет, чтобы завершить процесс вымирания.[5]
Человеческие воздействия
Подкисление
Диоксид серы и оксиды азота естественным образом выделяются из вулканов, органических соединений в почве, водно-болотных угодьях и морских системах, но большинство этих соединений возникает в результате сжигания угля, нефти, бензина и плавки руд, содержащих серу.[3] Эти вещества растворяются в атмосферной влаге и попадают в проточные системы в виде кислотный дождь.[1] Озера и пруды, содержащие коренные породы, богатые карбонатами, обладают естественным буфером, что не приводит к изменению pH. Однако системы без этой основной породы очень чувствительны к поступлению кислоты, поскольку они обладают низкой нейтрализующей способностью, что приводит к снижению pH даже при небольшом поступлении кислоты.[3] При pH 5–6 видовое разнообразие и биомасса водорослей значительно снижаются, что приводит к увеличению прозрачности воды - характерной особенности закисленных озер. По мере того, как pH продолжает снижаться, вся фауна становится менее разнообразной. Самая значимая особенность - нарушение воспроизводства рыб. Таким образом, популяция в конечном итоге состоит из нескольких старых особей, которые в конечном итоге умирают и оставляют системы без рыб.[2][3] Кислотные дожди особенно опасны для озер в Скандинавия, западный Шотландия, запад Уэльс и северо-восток Соединенных Штатов.
Эвтрофикация
Эвтрофный системы содержат высокую концентрацию фосфора (~ 30 мкг / л), азота (~ 1500 мкг / л) или того и другого.[2] Фосфор попадает в непроточные воды из очистка сточных вод сточные воды, сбросы из неочищенных сточных вод или с сельскохозяйственных угодий. Азот в основном поступает из удобрения для сельского хозяйства от стока или вымывания и последующего стока грунтовых вод. Это увеличение количества питательных веществ, необходимых для первичных продуцентов, приводит к значительному увеличению роста фитопланктона, что называется "цветение планктона. "Это цветение снижает прозрачность воды, что приводит к потере подводных растений. Результирующее сокращение структуры среды обитания отрицательно сказывается на видах, которые используют ее для нереста, созревания и общего выживания. Кроме того, большое количество недолговечного фитопланктона приводят к осаждению большого количества мертвой биомассы в отложениях.[5] Бактериям требуется большое количество кислорода для разложения этого материала, что снижает концентрацию кислорода в воде. Особенно ярко это выражено в стратифицированные озера, когда термоклин предотвращает смешивание с поверхности воды, богатой кислородом, с более низкими уровнями. Низкие или бескислородные условия исключают существование многих таксонов, которые физиологически не переносят эти условия.[2]
Инвазивные виды
Инвазивные виды были введены в проточные системы как в результате целенаправленных событий (например, зарыбление диких животных и кормовых видов), так и непреднамеренных событий (например, балластная вода ). Эти организмы могут влиять на аборигенов через конкуренцию за добычу или среду обитания, хищничество, изменение среды обитания, гибридизация, или занесение вредных болезней и паразитов.[4] Что касается местных видов, захватчики могут вызывать изменения в размерах и возрастной структуре, распределении, плотности, росте популяции и даже могут вести к исчезновению популяций.[2] Примеры известных захватчиков лентических систем включают зебра мидия и морская минога в Великих озерах.
Смотрите также
- Агентство по охране окружающей среды США - Экосистемы Великих озер
- Агентство по охране окружающей среды США - Учебник по лимнологии (файл PDF)
- Параметры качества окружающей среды пресной воды
- Лимнология
- Аэрация озера
- Искусственные непроточные водоемы Махараштры
использованная литература
- ^ а б c d е ж г час я j k л м п о п Браун, А. Л. (1987). Экология пресной воды. Образовательные книги Heinimann, Лондон. п. 163. ISBN 0435606220.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у Brönmark, C .; Л. А. Ханссон (2005). Биология озер и прудов. Издательство Оксфордского университета, Оксфорд. п. 285. ISBN 0198516134.
- ^ а б c d е ж г час я j k Калфф, Дж. (2002). Лимнология. Прентис Холл, Верхнее Седло, Нью-Джерси. п. 592. ISBN 0130337757.
- ^ а б c Giller, S .; Б. Мальмквист (1998). Биология ручьев и рек. Издательство Оксфордского университета, Оксфорд. п. 296. ISBN 0198549776.
- ^ а б c d е ж г Мосс, Б. (1998). Экология пресноводных водоемов: человек и среда, от прошлого к будущему. Blackwell Science, Лондон. п.557. ISBN 0632035129.
- ^ а б Кедди, П.А. (2010). Экология водно-болотных угодий: принципы и сохранение (2-е издание). Издательство Кембриджского университета, Кембридж, Великобритания. ISBN 0521739675.
- ^ а б Гливич, З. М. «Зоопланктон», стр. 461–516 в O'Sullivan (2005)
- ^ а б Йонассон, П. М. «Бентические беспозвоночные», стр. 341–416 в O'Sullivan (2005)
- ^ Winfield, I.J. "Экология популяции рыб", стр. 517–537 в O'Sullivan (2005)
- ^ Браун, Р. А. (1981). «Озера как острова: биогеографическое распространение, скорость оборота и видовой состав озер центральной части Нью-Йорка». Журнал биогеографии. 8 1: 75–83. Дои:10.2307/2844594. JSTOR 2844594.
- ^ Sommer, U .; З. М. Гливич; W. Lampert; А. Дункан (1986). «PEG-модель сезонной последовательности планктонных явлений в пресных водах». Archiv für Hydrobiologie. 106: 433–471.
- ^ Хиллебранд, Х. (2004). «Об общности градиента широтного разнообразия» (PDF). Американский натуралист. 163 (2): 192–211. Дои:10.1086/381004. PMID 14970922.
- ^ Hillebrand, H .; Азовский А.И. (2001). «Размер тела определяет силу градиента широтного разнообразия». Экография. 24 (3): 251–256. Дои:10.1034 / j.1600-0587.2001.240302.x.
Список используемой литературы
- О'Салливан, Патрик; Рейнольдс, С. С. (2005). Справочник по озерам: восстановление и реабилитация озер. Вайли. ISBN 978-0-632-04795-6.


| Пруды |
|  |
|---|---|---|
| Бассейны | ||
| Лужи | ||
| Биота | ||
| Экосистемы | ||
| Связанный | ||