Цитохром с оксидаза - Cytochrome c oxidase
| Цитохром с оксидаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 1.9.3.1 | ||||||||
| Количество CAS | 9001-16-5 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| Цитохром с оксидаза | |
|---|---|
 | |
| Идентификаторы | |
| Символ | Цитохром с оксидаза |
| OPM суперсемейство | 4 |
| Белок OPM | 2дыр |
| Мембранома | 257 |
В фермент цитохром с оксидаза или Комплекс IV, EC 1.9.3.1, это большой трансмембранный белок комплекс найден в бактерии, археи, а митохондрии из эукариоты.[1]
Это последний фермент в дыхательных путях. электронная транспортная цепь из клетки расположен в мембране. Он получает электрон от каждого из четырех цитохром с молекул, и передает их одной молекуле дикислорода, превращая молекулярный кислород до двух молекул воды. В этом процессе он связывает четыре протона из внутренней водной фазы, образуя две молекулы воды, и перемещает еще четыре протона через мембрану, увеличивая трансмембранную разницу протонов. электрохимический потенциал который АТФ-синтаза затем использует для синтеза АТФ.
Структура
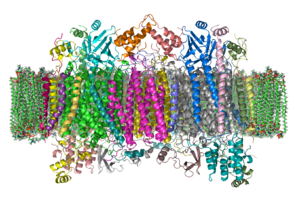
Комплекс
Комплекс большой интегральный мембранный белок состоит из нескольких металлические протезы и 14 [2] белковые субъединицы у млекопитающих. У млекопитающих одиннадцать субъединиц имеют ядерное происхождение, а три синтезируются в митохондриях. Комплекс состоит из двух гемы, а цитохром а и цитохром а3, и два медь центров, CuА и CuB центры.[3] Фактически, цитохром а3 и CuB образуют биядерный центр, который является местом восстановления кислорода. Цитохром с, который восстанавливается предыдущим компонентом дыхательной цепи (комплекс цитохрома bc1, комплекс III), стыкуется рядом с CuА биядерный центр и передает ему электрон, окисляясь обратно до цитохрома с, содержащего Fe3+. Восстановленная CuА биядерный центр теперь передает электрон на цитохром а, который, в свою очередь, передает электрон на цитохром а3-CuB биядерный центр. Два иона металла в этом биядерном центре находятся на расстоянии 4,5 Å друг от друга и координируют гидроксид-ион в полностью окисленном состоянии.
Кристаллографические исследования цитохром с оксидазы демонстрируют необычную посттрансляционную модификацию, связывающую C6 Tyr (244) и ε-N His (240) (нумерация бычьих ферментов). Он играет жизненно важную роль в обеспечении цитохрома а3- CuB двухъядерный центр, чтобы принять четыре электрона в восстановлении молекулярной кислород к воды. Ранее считалось, что механизм редукции включает перекись промежуточный, который, как полагали, приводит к супероксид производство. Однако принятый в настоящее время механизм включает быстрое четырехэлектронное восстановление с немедленным разрывом кислородно-кислородной связи, избегая любого промежуточного соединения, которое может образовывать супероксид.[4]:865–866
Консервированные субъединицы
| Нет. | Название подразделения | Человек белок | Описание белка из UniProt | Pfam семья с человеческим белком |
|---|---|---|---|---|
| 1 | Cox1 | COX1_HUMAN | Субъединица 1 цитохром с оксидазы | Pfam PF00115 |
| 2 | Cox2 | COX2_HUMAN | Субъединица 2 цитохром с оксидазы | Pfam PF02790, Pfam PF00116 |
| 3 | Cox3 | COX3_HUMAN | Субъединица 3 цитохром с оксидазы | Pfam PF00510 |
| 4 | Cox4i1 | COX41_HUMAN | Субъединица 4 цитохром с оксидазы, изоформа 1, митохондриальная | Pfam PF02936 |
| 5 | Cox4a2 | COX42_HUMAN | Субъединица 4 цитохром с оксидазы, изоформа 2, митохондриальная | Pfam PF02936 |
| 6 | Cox5a | COX5A_HUMAN | Субъединица 5А цитохром с оксидазы, митохондриальная | Pfam PF02284 |
| 7 | Cox5b | COX5B_HUMAN | Субъединица 5B цитохром с оксидазы, митохондриальная | Pfam PF01215 |
| 8 | Cox6a1 | CX6A1_HUMAN | Субъединица цитохром с оксидазы 6А1, митохондриальная | Pfam PF02046 |
| 9 | Cox6a2 | CX6A2_HUMAN | Субъединица цитохром с оксидазы 6A2, митохондриальная | Pfam PF02046 |
| 10 | Cox6b1 | CX6B1_HUMAN | Субъединица цитохром с оксидазы 6B1 | Pfam PF02297 |
| 11 | Cox6b2 | CX6B2_HUMAN | Субъединица цитохром с оксидазы 6B2 | Pfam PF02297 |
| 12 | Cox6c | COX6C_HUMAN | Субъединица 6C цитохром с оксидазы | Pfam PF02937 |
| 13 | Cox7a1 | CX7A1_HUMAN | Субъединица 7A1 цитохром с оксидазы, митохондриальная | Pfam PF02238 |
| 14 | Cox7a2 | CX7A2_HUMAN | Субъединица 7A2 цитохром с оксидазы, митохондриальная | Pfam PF02238 |
| 15 | Cox7a3 | COX7S_HUMAN | Предполагаемая субъединица цитохром с оксидазы 7A3, митохондриальная | Pfam PF02238 |
| 16 | Cox7b | COX7B_HUMAN | Субъединица 7B цитохром с оксидазы, митохондриальная | Pfam PF05392 |
| 17 | Cox7c | COX7C_HUMAN | Субъединица 7C цитохром с оксидазы, митохондриальная | Pfam PF02935 |
| 18 | Cox7r | COX7R_HUMAN | Белок, связанный с субъединицей 7А цитохром с оксидазы, митохондриальный | Pfam PF02238 |
| 19 | Cox8a | COX8A_HUMAN | Субъединица цитохром с оксидазы 8A, митохондриальный P | Pfam PF02285 |
| 20 | Cox8c | COX8C_HUMAN | Субъединица цитохром с оксидазы 8C, митохондриальная | Pfam PF02285 |
| Подразделения сборки[7][8][9] | ||||
| 1 | Coa1 | COA1_HUMAN | Гомолог фактора сборки 1 цитохром с оксидазы | Pfam PF08695 |
| 2 | Coa3 | COA3_HUMAN | Гомолог фактора сборки цитохром с оксидазы 3, митохондриальный | Pfam PF09813 |
| 3 | Coa4 | COA4_HUMAN | Гомолог фактора сборки 4 цитохром с оксидазы, митохондриальный | Pfam PF06747 |
| 4 | Coa5 | COA5_HUMAN | Фактор сборки цитохром с оксидазы 5 | Pfam PF10203 |
| 5 | Coa6 | COA6_HUMAN | Гомолог фактора сборки цитохром с оксидазы 6 | Pfam PF02297 |
| 6 | Coa7 | COA7_HUMAN | Фактор сборки цитохром с оксидазы 7, | Pfam PF08238 |
| 7 | Cox11 | COX11_HUMAN | Белок сборки цитохром-с-оксидазы COX11 митохондрии | Pfam PF04442 |
| 8 | Cox14 | COX14_HUMAN | Сборочный белок цитохром с оксидазы | Pfam PF14880 |
| 9 | Cox15 | COX15_HUMAN | Гомолог белка сборки цитохром с оксидазы COX15 | Pfam PF02628 |
| 10 | Cox16 | COX16_HUMAN | Гомолог митохондриального белка сборки цитохром с оксидазы COX16 | Pfam PF14138 |
| 11 | Cox17 | COX17_HUMAN | Цитохром с оксидаза медный шаперон | Pfam PF05051 |
| 12 | Cox18[10] | COX18_HUMAN | Белок внутренней мембраны митохондрий (сборочный белок цитохром с оксидазы 18) | Pfam PF02096 |
| 13 | Cox19 | COX19_HUMAN | Сборочный белок цитохром с оксидазы | Pfam PF06747 |
| 14 | Cox20 | COX20_HUMAN | Гомолог белка цитохром с оксидазы 20 | Pfam PF12597 |
Сборка
Сборка СОХ в дрожжи представляет собой сложный процесс, который не совсем понятен из-за быстрой и необратимой агрегации гидрофобных субъединиц, которые образуют холоферментный комплекс, а также агрегации мутантных субъединиц с открытыми гидрофобными участками.[11] Субъединицы СОХ кодируются как в ядерном, так и в митохондриальном геномах. Три субъединицы, образующие каталитическое ядро ЦОГ, кодируются в митохондриальном геноме.
Гемы и кофакторы вставлены в субъединицы I и II. Две молекулы гема находятся в субъединице I, помогая транспорту к субъединице II, где две молекулы меди способствуют непрерывному переносу электронов.[12] Подразделения I и IV инициируют сборку. Различные субъединицы могут связываться с образованием промежуточных субкомплексов, которые позже связываются с другими субъединицами с образованием комплекса СОХ.[11] В модификациях после сборки ЦОГ образует гомодимер. Это необходимо для активности. Оба димера связаны между собой кардиолипин молекула[11][13][14] который, как было обнаружено, играет ключевую роль в стабилизации холоферментного комплекса. Диссоциация субъединиц VIIa и III в сочетании с удалением кардиолипина приводит к полной потере активности фермента.[14] Известно, что субъединицы, кодируемые в ядерном геноме, играют роль в димеризации и стабильности ферментов. Мутации в этих субъединицах устраняют функцию ЦОГ.[11]
Известно, что сборка происходит по крайней мере за три различных этапа, определяющих скорость. Были обнаружены продукты этих стадий, хотя состав конкретных субъединиц не был определен.[11]
Синтезу и сборке субъединиц ЦОГ I, II и III способствуют активаторы трансляции, которые взаимодействуют с 5 ’нетранслируемыми областями транскриптов митохондриальной мРНК. Активаторы трансляции кодируются в ядре. Они могут действовать посредством прямого или косвенного взаимодействия с другими компонентами машинного перевода, но точные молекулярные механизмы неясны из-за трудностей, связанных с синтезом машинного перевода in-vitro.[15][16] Хотя взаимодействия между субъединицами I, II и III, кодируемые в митохондриальном геноме, вносят меньший вклад в стабильность фермента, чем взаимодействия между бигеномными субъединицами, эти субъединицы более консервативны, что указывает на потенциальную неизученную роль для активности фермента.[17]
Биохимия
Суммарная реакция:
- 4 Fe2+-цитохром c + 8 часов+в + O2 → 4 Fe3+-цитохром c + 2 часа2O + 4 H+вне
Два электрона проходят от двух цитохромов с через CuА и цитохром а сайты цитохрома а3- CuB двухъядерный центр, восстанавливающий металлы до Fe2+ форма и Cu+. Гидроксидный лиганд протонируется и теряется в виде воды, создавая пустоту между металлами, которая заполняется O2. Кислород быстро восстанавливается, и два электрона поступают из Fe2+цитохром а3, который превращается в феррилоксоформу (Fe4+= O). Атом кислорода, близкий к CuB забирает один электрон от Cu+, а второй электрон и протон из гидроксил Tyr (244), который становится тирозильным радикалом. Второй кислород превращается в гидроксид-ион, собирая два электрона и протон. Третий электрон, возникающий из другого цитохрома с, передается через первые два переносчика электронов цитохрому а.3- CuB двухъядерный центр, и этот электрон и два протона превращают тирозильный радикал обратно в Tyr, а гидроксид, связанный с CuB2+ к молекуле воды. Четвертый электрон от другого цитохрома с проходит через CuА и цитохром а к цитохрому а3- CuB биядерный центр, восстанавливающий Fe4+= O в Fe3+, причем атом кислорода одновременно захватывает протон, регенерируя этот кислород в виде гидроксид-иона, координированного в середине цитохрома а.3- CuB центр, как это было в начале этого цикла. Чистый процесс состоит в том, что четыре восстановленных цитохрома с используются вместе с 4 протонами для восстановления O2 до двух молекул воды.[4]:841–5
Торможение
ЦОГ существует в трех конформационных состояниях: полностью окисленный (импульсный), частично восстановленный и полностью восстановленный. Каждый ингибитор имеет высокое сродство к разному состоянию. В импульсном состоянии оба гема a3 и CuB ядерные центры окисляются; это конформация фермента, имеющего наивысшую активность. Двухэлектронное восстановление инициирует конформационное изменение, которое позволяет кислороду связываться в активном центре с частично восстановленным ферментом. Четыре электрона связываются с ЦОГ, чтобы полностью восстановить фермент. Его полностью восстановленное состояние, которое состоит из восстановленного Fe2+ на цитохроме а3 гемовая группа и восстановленная CuB+ биядерный центр считается неактивным или состоянием покоя фермента.[18]
Цианид, азид, и монооксид углерода[19] все связываются с цитохром с оксидазой, препятствуя функционированию белка и приводя к химическому удушье ячеек. Более высокие концентрации молекулярного кислорода необходимы для компенсации возрастающих концентраций ингибитора, что приводит к общему снижению метаболической активности в клетке в присутствии ингибитора. Другие лиганды, такие как оксид азота и сероводород, также могут ингибировать ЦОГ, связываясь с регуляторными участками фермента, снижая скорость клеточного дыхания.[20]
Цианид - неконкурентный ингибитор ЦОГ,[21][22] связывание с высоким сродством с частично восстановленным состоянием фермента и препятствие дальнейшему восстановлению фермента. В импульсном состоянии цианид связывается медленно, но с высоким сродством. Лиганд предназначен для электростатической стабилизации обоих металлов одновременно, располагаясь между ними. Высокая концентрация оксида азота, например, экзогенно добавляемая к ферменту, отменяет цианидное ингибирование ЦОГ.[23]
Оксид азота может обратимо[24] связываются с любым ионом металла в биядерном центре, чтобы окислиться до нитрита. NO и CN− будет конкурировать с кислородом за связывание на этом участке, снижая скорость клеточного дыхания. Однако эндогенный NO, который продуцируется на более низких уровнях, увеличивает CN.− торможение. Более высокие уровни NO, которые коррелируют с наличием большего количества фермента в восстановленном состоянии, приводят к большему ингибированию цианида.[18] Известно, что при этих базовых концентрациях NO ингибирование Комплекса IV имеет положительные эффекты, такие как повышение уровня кислорода в тканях кровеносных сосудов. Неспособность фермента преобразовывать кислород в воду приводит к накоплению кислорода, который может глубже проникать в окружающие ткани.[24] NO-ингибирование Комплекса IV имеет больший эффект при более низких концентрациях кислорода, увеличивая его полезность в качестве сосудорасширяющего средства в тканях, где это необходимо.[24]
Сероводород будет связывать ЦОГ неконкурентным образом на регуляторном участке фермента, подобно монооксиду углерода. Сульфид имеет наивысшее сродство к импульсному или частично восстановленному состоянию фермента и способен частично восстанавливать фермент на геме а.3 центр. Неясно, является ли эндогенный H2Уровни S достаточны для подавления фермента. Нет взаимодействия между сероводородом и полностью восстановленной конформацией СОХ.[20]
Метанол в денатурат превращается в Муравьиная кислота, который также ингибирует ту же оксидазную систему. Высокий уровень АТФ может аллостерически ингибируют цитохром-с-оксидазу, связываясь внутри митохондриального матрикса.[25]
Внемитохондриальная и субклеточная локализации

Цитохром с оксидаза состоит из 3 субъединиц, которые кодируются митохондриальная ДНК (цитохром с оксидаза субъединица I, субъединица II, и субъединица III ). Из этих 3 субъединиц, кодируемых митохондриальной ДНК, две были идентифицированы во внемитохондриальных местах. В панкреатический ацинарной ткани эти субъединицы были обнаружены в зимоген гранулы. Кроме того, в передний гипофиз, относительно высокие количества этих субъединиц были обнаружены в гормон роста секреторные гранулы.[26] Экстрамитохондриальная функция этих субъединиц цитохром с оксидазы еще не охарактеризована. Помимо субъединиц цитохром с оксидазы, внемитохондриальная локализация также наблюдалась для большого количества других митохондриальных белков.[27][28] Это поднимает возможность существования пока неустановленных специфических механизмов транслокации белков из митохондрий в другие клеточные места назначения.[26][28][29]
Генетические дефекты и нарушения
Дефекты, связанные с генетическими мутациями, изменяющими цитохром c функциональность или структура оксидазы (СОХ) может привести к тяжелым, часто смертельным метаболические нарушения. Такие нарушения обычно проявляются в раннем детстве и поражают преимущественно ткани с высокими потребностями в энергии (мозг, сердце, мышцы). Среди множества засекреченных митохондриальные заболевания те, которые связаны с дисфункциональной сборкой ЦОГ, считаются наиболее тяжелыми.[30]
Подавляющее большинство нарушений ЦОГ связаны с мутациями в ядерно-кодируемых белках, которые называются факторами сборки или сборочными белками. Эти факторы сборки вносят вклад в структуру и функциональность СОХ и участвуют в нескольких важных процессах, включая транскрипцию и трансляцию субъединиц, кодируемых митохондриями, процессинг препротеинов и вставку в мембрану, а также биосинтез и включение кофакторов.[31]
В настоящее время мутации идентифицированы в семи факторах сборки ЦОГ: SURF1, SCO1, SCO2, COX10, COX15, COX20, COA5 и LRPPRC. Мутации в этих белках могут привести к изменению функциональности сборки субкомплекса, транспорта меди или регуляции трансляции. Каждая мутация гена связана с этиологией конкретного заболевания, при этом некоторые из них имеют последствия для множества заболеваний. Заболевания, связанные с дисфункциональной сборкой ЦОГ посредством генных мутаций, включают: Синдром Ли, кардиомиопатия, лейкодистрофия, анемия, и нейросенсорная глухота.
Гистохимия
Повышенная зависимость нейронов от окислительного фосфорилирования для получения энергии[32] облегчает использование гистохимии ЦОГ для картирования регионального метаболизма головного мозга у животных, поскольку устанавливает прямую и положительную корреляцию между активностью ферментов и нейронами.[33] Это можно увидеть по корреляции между количеством и активностью фермента ЦОГ, что указывает на регуляцию ЦОГ на уровне экспрессии генов. Распределение ЦОГ непостоянно в разных областях мозга животных, но характер его распределения у животных одинаков. Эта закономерность наблюдалась в мозге обезьяны, мыши и теленка. Один изофермент ЦОГ постоянно обнаруживается при гистохимическом анализе мозга.[34]
Такое картирование мозга было выполнено у спонтанных мутантных мышей с заболеванием мозжечка, таким как моталка[35] и трансгенная модель Болезнь Альцгеймера.[36] Этот метод также использовался для составления карты учебной активности в мозгу животных.[37]
Дополнительные изображения

ТАК ДАЛЕЕ

Комплекс IV

Смотрите также
- Субъединица I цитохром с оксидазы
- Субъединица II цитохром с оксидазы
- Субъединица III цитохром с оксидазы
- Heme a
использованная литература
- ^ Кастресана Дж., Люббен М., Сарасте М., Хиггинс Д. Г. (июнь 1994 г.). «Эволюция цитохромоксидазы, фермента старше атмосферного кислорода». Журнал EMBO. 13 (11): 2516–2525. Дои:10.1002 / j.1460-2075.1994.tb06541.x. ЧВК 395125. PMID 8013452.
- ^ Бальса Е., Марко Р., Пералес-Клементе Е., Шкларчик Р., Кальво Е., Ландазури М. О., Энрикес Дж. А. (сентябрь 2012 г.). «NDUFA4 является субъединицей комплекса IV электронно-транспортной цепи млекопитающих». Клеточный метаболизм. 16 (3): 378–86. Дои:10.1016 / j.cmet.2012.07.015. PMID 22902835.
- ^ Цукихара Т., Аояма Х., Ямасита Э., Томизаки Т., Ямагути Х., Синдзава-Ито К., Накашима Р., Яоно Р., Йошикава С. (август 1995 г.). «Структуры металлических сайтов окисленной цитохром с оксидазы сердца крупного рогатого скота при 2,8 А». Наука. 269 (5227): 1069–74. Bibcode:1995Научный ... 269.1069T. Дои:10.1126 / science.7652554. PMID 7652554. S2CID 27210776.
- ^ а б Воет Д., Воет Дж. Г. (2011). Биохимия (4-е изд.). Хобокен, Нью-Джерси: Джон Уайли и сыновья. ISBN 978-0-470-57095-1.
- ^ Чжан З., Хуанг Л., Шульмейстер В.М., Чи Ю.И., Ким К.К., Хунг Л.В., Крофтс А.Р., Берри Е.А., Ким С.Х. (апрель 1998 г.). «Перенос электронов путем перемещения домена в цитохроме bc1». Природа. 392 (6677): 677–84. Bibcode:1998Натура.392..677Z. Дои:10.1038/33612. PMID 9565029. S2CID 4380033.
- ^ Кайла В.Р., Оксанен Э., Гольдман А., Блох Д.А., Верховский М.И., Сундхольм Д., Викстрём М. (июль 2011 г.). «Комбинированное квантово-химическое и кристаллографическое исследование окисленного биядерного центра цитохром с оксидазы». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1807 (7): 769–78. Дои:10.1016 / j.bbabio.2010.12.016. PMID 21211513.
- ^ Szklarczyk R, Wanschers BF, Cuypers TD, Esseling JJ, Riemersma M, van den Brand MA, Gloerich J, Lasonder E, van den Heuvel LP, Nijtmans LG, Huynen MA (февраль 2012 г.). «Итеративное прогнозирование ортологии обнаруживает новые митохондриальные белки и идентифицирует C12orf62 как человеческий ортолог COX14, белка, участвующего в сборке цитохром с оксидазы». Геномная биология. 13 (2): R12. Дои:10.1186 / gb-2012-13-2-r12. ЧВК 3334569. PMID 22356826.
- ^ Mick DU, Dennerlein S, Wiese H, Reinhold R, Pacheu-Grau D, Lorenzi I, Sasarman F, Weraarpachai W., Shoubridge EA, Warscheid B, Rehling P (декабрь 2012 г.). «MITRAC связывает транслокацию митохондриальных белков со сборкой дыхательной цепи и регуляцией трансляции». Ячейка. 151 (7): 1528–41. Дои:10.1016 / j.cell.2012.11.053. PMID 23260140.
- ^ Козьяк-Павлович В., Прелл Ф., Тиде Б., Гётц М., Возиек Д., Отт С., Рудель Т. (февраль 2014 г.). «C1orf163 / RESA1 - это новый белок митохондриального межмембранного пространства, связанный со сборкой дыхательной цепи». Журнал молекулярной биологии. 426 (4): 908–20. Дои:10.1016 / j.jmb.2013.12.001. PMID 24333015.
- ^ Gaisne M, Bonnefoy N (сентябрь 2006 г.). «Ген СОХ18, участвующий в митохондриальном биогенезе, функционально консервативен и строго регулируется у людей и делящихся дрожжей». FEMS дрожжевые исследования. 6 (6): 869–82. Дои:10.1111 / j.1567-1364.2006.00083.x. PMID 16911509.
- ^ а б c d е Fontanesi F, Soto IC, Horn D, Barrientos A (декабрь 2006 г.). «Сборка митохондриальной цитохром с-оксидазы, сложный и строго регулируемый клеточный процесс». Американский журнал физиологии. Клеточная физиология. 291 (6): C1129-47. Дои:10.1152 / ajpcell.00233.2006. PMID 16760263.
- ^ Крофтс А (1996). «Цитохромоксидаза: Комплекс IV». Университет Иллинойса в Урбане-Шампейн.
- ^ Халимончук О., Рёдель Г. (декабрь 2005 г.). «Биогенез цитохром с оксидазы». Митохондрия. 5 (6): 363–88. Дои:10.1016 / j.mito.2005.08.002. PMID 16199211.
- ^ а б Седлак Э., Робинсон NC (сентябрь 2015 г.). «Дестабилизация четвертичной структуры цитохром с оксидазы сердца крупного рогатого скота при удалении прочно связанного кардиолипина». Биохимия. 54 (36): 5569–77. Дои:10.1021 / acs.biochem.5b00540. PMID 26284624.
- ^ Херрманн Дж. М., Вельхаф М. В., Боннефой Н. (февраль 2013 г.). «Контроль синтеза белка в митохондриях дрожжей: концепция активаторов трансляции». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1833 (2): 286–94. Дои:10.1016 / j.bbamcr.2012.03.007. PMID 22450032.
- ^ Сото И.К., Фонтанези Ф., Лю Дж., Барриентос А. (июнь 2012 г.). «Биогенез и сборка каталитического ядра цитохром с оксидазы эукариот». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1817 (6): 883–97. Дои:10.1016 / j.bbabio.2011.09.005. ЧВК 3262112. PMID 21958598.
- ^ Аледо Х.С., Вальверде Х., Руис-Камачо М., Морилла I, Лопес ФЗ (октябрь 2014 г.). «Интерфейсы белок-белок цитохром с оксидазы I развиваются быстрее, чем несвязывающие поверхности, но отрицательный отбор является движущей силой». Геномная биология и эволюция. 6 (11): 3064–76. Дои:10.1093 / gbe / evu240. ЧВК 4255772. PMID 25359921.
- ^ а б Ливсли Х. Б., Ли Л., Прабхакаран К., Боровиц Дж. Л., Изом Г. Е. (январь 2008 г.). «Взаимодействие цианида и оксида азота с цитохром с оксидазой: последствия для острой токсичности цианида». Токсикологические науки. 101 (1): 101–11. Дои:10.1093 / toxsci / kfm254. PMID 17906319.
- ^ Алонсо Дж. Р., Карделлах Ф., Лопес С., Касадемонт Дж., Миро О. (сентябрь 2003 г.). «Окись углерода специфически ингибирует цитохром-с-оксидазу митохондриальной дыхательной цепи человека». Фармакология и токсикология. 93 (3): 142–6. Дои:10.1034 / j.1600-0773.2003.930306.x. PMID 12969439.
- ^ а б Николс П., Маршалл, округ Колумбия, Купер CE, Уилсон MT (октябрь 2013 г.). «Сульфидное ингибирование и метаболизм цитохром с оксидазой». Сделки Биохимического Общества. 41 (5): 1312–6. Дои:10.1042 / BST20130070. PMID 24059525. S2CID 11554252.
- ^ Робертс М., Рейсс М.Дж., Монгер Г. (2000). Продвинутая биология. Нельсон Торнс. ISBN 9780174387329.
- ^ Робертс МБ (1986). Биология: функциональный подход. Нельсон Торнс. ISBN 9780174480198.
- ^ Дженсен П., Уилсон М. Т., Ааса Р., Мальмстрем Б. Г. (декабрь 1984 г.). «Цианидное ингибирование цитохром-с-оксидазы. Исследование e.p.r. с быстрым замораживанием». Биохимический журнал. 224 (3): 829–37. Дои:10.1042 / bj2240829. ЧВК 1144519. PMID 6098268.
- ^ а б c Гладвин М. Т., Шива С. (май 2009 г.). «Битва за связывание лигандов в цитохром с оксидазе: как NO регулирует градиенты кислорода в тканях». Циркуляционные исследования. 104 (10): 1136–8. Дои:10.1161 / CIRCRESAHA.109.198911. PMID 19461104.
- ^ Арнольд С., Каденбах Б. (октябрь 1997 г.). «Дыхание клеток контролируется АТФ, аллостерическим ингибитором цитохром-с оксидазы». Eur J Biochem. 249 (1): 350–354. Дои:10.1111 / j.1432-1033.1997.t01-1-00350.x. PMID 9363790.
- ^ а б Садачаран С.К., Сингх Б., Боуз Т., Гупта Р.С. (ноябрь 2005 г.). «Локализация митохондриальной ДНК, кодируемой субъединицами I и II цитохром с оксидазы в гранулах зимогена поджелудочной железы крысы и гранулах гормона роста гипофиза». Гистохимия и клеточная биология. 124 (5): 409–21. Дои:10.1007 / s00418-005-0056-2. PMID 16133117. S2CID 24440427.
- ^ Гупта Р.С., Рамачандра Н.Б., Боуз Т., Сингх Б. (2008). «Необычное расположение в клетках митохондриальных молекулярных шаперонов Hsp60, Hsp70 и Hsp10». Симпозиум Фонда Новартис. Симпозиумы Фонда Новартис. 291: 59–68, обсуждение 69–73, 137–40. Дои:10.1002 / 9780470754030.ch5. ISBN 9780470754030. PMID 18575266.
- ^ а б Солтыс Б.Дж., Гупта Р.С. (1999). «Митохондриальные белки в неожиданных клеточных местоположениях: экспорт белков из митохондрий с эволюционной точки зрения». Международный обзор цитологии. 194: 133–96. Дои:10.1016 / S0074-7696 (08) 62396-7. ISBN 9780123645982. PMID 10494626.
- ^ Солтыс Б.Дж., Гупта Р.С. (май 1999 г.). «Белки митохондриального матрикса в неожиданных местах: экспортируются ли они?». Тенденции в биохимических науках. 24 (5): 174–7. Дои:10.1016 / s0968-0004 (99) 01390-0. PMID 10322429.
- ^ Pecina P, Houstková H, Hansíková H, Zeman J, Houstek J (2004). «Генетические дефекты сборки цитохром с оксидазы» (PDF). Физиологические исследования. 53 Приложение 1: S213-23. PMID 15119951.
- ^ Zee JM, Glerum DM (декабрь 2006 г.). «Дефекты сборки цитохромоксидазы у человека: уроки дрожжей». Биохимия и клеточная биология. 84 (6): 859–69. Дои:10.1139 / o06-201. PMID 17215873.
- ^ Джохар К., Прия А., Дхар С., Лю К., Вонг-Райли М. Т. (ноябрь 2013 г.). «Нейрон-специфичный белок 4 бигеномно регулирует транскрипцию всех генов субъединиц цитохром с оксидазы, кодируемых митохондриями и ядром, в нейронах». Журнал нейрохимии. 127 (4): 496–508. Дои:10.1111 / jnc.12433. ЧВК 3820366. PMID 24032355.
- ^ Вонг-Райли MT (март 1989 г.). «Цитохромоксидаза: эндогенный метаболический маркер нейрональной активности». Тенденции в неврологии. 12 (3): 94–101. Дои:10.1016/0166-2236(89)90165-3. PMID 2469224. S2CID 42996304.
- ^ Хевнер РФ, Вонг-Райли MT (ноябрь 1989 г.). «Цитохромоксидаза головного мозга: очистка, выработка антител и иммуногистохимические / гистохимические корреляции в ЦНС». Журнал неврологии. 9 (11): 3884–98. Дои:10.1523 / jneurosci.09-11-03884.1989. ЧВК 6569932. PMID 2555458.
- ^ Strazielle C, Hayzoun K, Derer M, Mariani J, Lalonde R (апрель 2006 г.). «Региональные мозговые вариации активности цитохромоксидазы у мышей с мутантом Relnrl-orl». Журнал неврологических исследований. 83 (5): 821–31. Дои:10.1002 / jnr.20772. PMID 16511878. S2CID 45787322.
- ^ Strazielle C, Sturchler-Pierrat C, Staufenbiel M, Lalonde R (2003). «Региональная активность цитохромоксидазы головного мозга у трансгенных мышей бета-амилоидного белка-предшественника с мутацией Swedish». Неврология. 118 (4): 1151–63. Дои:10.1016 / S0306-4522 (03) 00037-X. PMID 12732258. S2CID 9366458.
- ^ Конехо Н.М., Гонсалес-Пардо Х., Гонсалес-Лима Ф., Ариас Дж. Л. (март 2010 г.). «Пространственное обучение водного лабиринта: прогрессия мозговых цепей, отображенных с помощью гистохимии цитохромоксидазы». Нейробиология обучения и памяти. 93 (3): 362–71. Дои:10.1016 / j.nlm.2009.12.002. PMID 19969098. S2CID 24271956.
внешние ссылки
- Домашняя страница цитохромоксидазы в Университет Райса
- Интерактивная молекулярная модель цитохром с оксидазы (Требуется MDL Chime )
- UMich Ориентация белков в мембранах семьи / суперсемейство-4
- Цитохром-с + оксидаза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)