Клональный отбор - Clonal selection
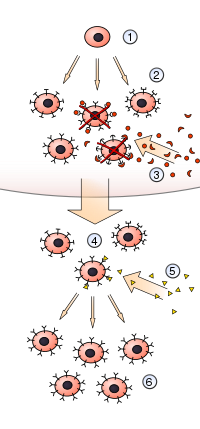
Теория клонального отбора это научная теория в иммунология что объясняет функции клеток иммунная система (лимфоциты ) в ответ на конкретные антигены вторжение в тело. Концепция была представлена австралийским врачом Фрэнк Макфарлейн Бернет в 1957 г., пытаясь объяснить большое разнообразие антитела сформированный во время инициирования иммунная реакция.[1][2] Теория стала общепринятой моделью того, как человек иммунная система отвечает на инфекционное заболевание и как определенные типы B и Т-лимфоциты отбираются для уничтожения конкретных антигены.[3]
Теория утверждает, что в ранее существовавшей группе лимфоцитов (в частности, В-лимфоцитах) определенный антиген активирует (то есть отбирает) только свою контрспецифичную клетку, которая затем побуждает эту конкретную клетку размножаться, производя идентичные клоны для производства антител. Эта активация происходит во вторичных лимфоидных органах, таких как селезенка и лимфатический узел.[4] Короче говоря, теория - это объяснение механизма создания разнообразия специфичности антител.[5] Первые экспериментальные доказательства были получены в 1958 году, когда Густав Носсаль и Джошуа Ледерберг показали, что одна В-клетка всегда производит только одно антитело.[6] Идея оказалась основой молекулярной иммунологии, особенно в адаптивный иммунитет.[7]
Постулаты
Теорию клонального отбора можно резюмировать следующими четырьмя принципами:
- Каждый лимфоцит несет один тип рецептора с уникальной специфичностью (генерируемый V (D) J рекомбинация ).
- Для активации клеток требуется оккупация рецептора.
- Дифференцированный эффекторные клетки полученные из активированных лимфоцитов несут рецепторы, обладающие такой же специфичностью, как и родительская клетка.
- Те лимфоциты, несущие рецепторы собственных молекул (т. Е. эндогенный антигены, продуцируемые в организме) разрушаются на ранней стадии.
Ранняя работа
В 1900 г. Пол Эрлих предложил так называемую «теорию боковой цепи» производства антител. Согласно ему, определенные клетки проявляют на своей поверхности разные "боковые цепи «(т.е. мембраносвязанные антитела), способные реагировать с различными антигенами. Когда антиген присутствует, он связывается с соответствующей боковой цепью. Затем клетка перестает продуцировать все другие боковые цепи и начинает интенсивный синтез и секрецию антигенсвязывающей стороны В качестве растворимого антитела идея Эрлиха, хотя и отличается от клональной селекции, была теорией селекции, гораздо более точной, чем поучительные теории, доминировавшие в иммунологии в следующие десятилетия.
В 1955 году датский иммунолог. Нильс Йерн выдвинули гипотезу о том, что уже существует огромное количество растворимых антител в сыворотка до заражения. Попадание антигена в организм приводит к отбору подходящего антитела только одного типа. Это предположительно произошло некоторыми клетками фагоцитозирование иммунные комплексы и каким-то образом реплицируют структуру антител, чтобы производить их больше.[8]
В 1957 г. Дэвид В. Талмейдж выдвинули гипотезу, что антигены связываются с антителами на поверхности продуцирующих антитела клеток и «для размножения отбираются только те клетки, синтезированный продукт которых имеет сродство к антигену». Ключевым отличием от теории Эрлиха было то, что предполагалось, что каждая клетка синтезирует только один вид антител. После связывания антигена клетка размножается, образуя клоны с идентичными антителами.
Теория клонального отбора Бернета
Позже в 1957 году австралийский иммунолог Фрэнк Макфарлейн Бернет опубликовал статью под названием «Модификация теории продуцирования антител Джерна с использованием концепции клонального отбора» в довольно малоизвестной Австралийский научный журнал. В ней Бернет расширил идеи Талмейджа и назвал получившуюся теорию «теорией клонального отбора». Он далее формализовал теорию в своей книге 1959 года. Теория клонального отбора приобретенного иммунитета. Он объяснил иммунологическая память как клонирование двух типов лимфоцитов. Один клон действует немедленно, чтобы бороться с инфекцией, в то время как другой действует дольше, оставаясь в иммунной системе в течение длительного времени и вызывая иммунитет к этому антигену. Согласно гипотезе Бернета, среди антител есть молекулы, которые, вероятно, могут с разной степенью точности соответствовать всем или практически всем, антигенные детерминанты которые происходят в биологическом материале, отличном от тех, которые характерны для самого тела. Каждый тип паттерна является конкретным продуктом клона лимфоцитов, и это суть гипотезы о том, что каждая клетка автоматически имеет на своей поверхности репрезентативные реактивные участки, эквивалентные участкам клетки. глобулин они производят. Когда антиген попадает в кровь или тканевые жидкости, предполагается, что он будет прикрепляться к поверхности любого лимфоцита, несущего реактивные участки, соответствующие одной из его антигенных детерминант. Затем клетка активируется и подвергается пролиферации, чтобы произвести множество потомков. Таким образом, инициируется преимущественная пролиферация всех тех клонов, реактивные сайты которых соответствуют антигенным детерминантам антигенов, присутствующих в организме. Потомки способны к активному высвобождению растворимых антител и лимфоцитов, выполняя те же функции, что и родительские формы.[5][9]
В 1958 г. Густав Носсаль и Джошуа Ледерберг показали, что одна В-клетка всегда продуцирует только одно антитело, что было первым прямым доказательством, подтверждающим теорию клональной селекции.[6]
Теории, поддерживаемые клональным отбором
Бернет и Питер Медавар работали вместе над пониманием иммунологическая толерантность, явление также объясняется клональным отбором. Это способность организма переносить введение клеток до развития иммунного ответа, если это происходит на ранней стадии развития организма. В иммунной системе встречается огромное количество лимфоцитов, от клеток, которые переносят собственную ткань, до клеток, которые этого не делают. Однако только клетки, толерантные к собственной ткани, выживают на эмбриональной стадии. При введении чужеродной ткани развиваются лимфоциты, которые включают чужеродные ткани как собственную ткань.
В 1959 году Бернет предположил, что при определенных обстоятельствах ткани могут быть успешно трансплантированы иностранным реципиентам. Эта работа привела к гораздо большему пониманию иммунной системы, а также к большим достижениям в трансплантации тканей. Бернет и Медавар разделили Нобелевская премия по физиологии и медицине в 1960 г.
В 1974 г. Нильс Кай Йерн предположили, что иммунная система функционирует как сеть, которая регулируется посредством взаимодействий между вариабельными частями лимфоцитов и их секретируемыми молекулами. Теория иммунных сетей прочно основывается на концепции клонального отбора. Джерн получил Нобелевскую премию по физиологии и медицине в 1984 году, в основном за его вклад в теорию иммунных сетей.
Смотрите также
Рекомендации
- ^ Бернет, FM (1976). «Модификация теории производства антител Джерна с использованием концепции клонального отбора». КА: Журнал онкологических заболеваний для клиницистов. 26 (2): 119–21. Дои:10.3322 / canjclin.26.2.119. PMID 816431. S2CID 40609269.
- ^ Кон, Мелвин; Av Mitchison, N .; Пол, Уильям Э .; Сильверштейн, Артур М .; Талмейдж, Дэвид В .; Вейгерт, Мартин (2007). «Размышления о теории клонального отбора». Nature Reviews Иммунология. 7 (10): 823–830. Дои:10.1038 / nri2177. PMID 17893695. S2CID 24741671.
- ^ Раевский, Клаус (1996). «Клональный отбор и обучение в системе антител». Природа. 381 (6585): 751–758. Дои:10.1038 / 381751a0. ISSN 0028-0836. PMID 8657279. S2CID 4279640.
- ^ Мерфи, Кеннет (2012). Иммунобиология Джейнвей, 8-е издание. Нью-Йорк, Нью-Йорк: Наука о гирляндах. ISBN 9780815342434.
- ^ а б Джордан, Маргарет А; Бакстер, Алан Дж. (2007). «Количественный и качественный подходы к БОГУ: первые 10 лет теории клонального отбора». Иммунология и клеточная биология. 86 (1): 72–79. Дои:10.1038 / sj.icb.7100140. PMID 18040281. S2CID 19122290.
- ^ а б Nossal, G. J. V .; Ледерберг, Джошуа (1958). «Продукция антител отдельными клетками». Природа. 181 (4620): 1419–1420. Дои:10.1038 / 1811419a0. ЧВК 2082245. PMID 13552693.
- ^ Меджитов, Р. (2013). «Теория распознавания образов и начало современного врожденного иммунитета». Журнал иммунологии. 191 (9): 4473–4474. Дои:10.4049 / jimmunol.1302427. PMID 24141853.
- ^ Бернет, Ф. М. (1976). «Модификация теории производства антител Джерна с использованием концепции клонального отбора». КА: Журнал онкологических заболеваний для клиницистов. 26 (2): 119–121. Дои:10.3322 / canjclin.26.2.119. ISSN 0007-9235. PMID 816431. S2CID 40609269.
- ^ Ходжкин, Филип Д; Хит, Уильям Р.; Бакстер, Алан Дж. (2007). «Теория клонального отбора: 50 лет революции». Иммунология природы. 8 (10): 1019–1026. Дои:10.1038 / ni1007-1019. PMID 17878907. S2CID 29935594.
дальнейшее чтение
- Подольский, Альфред И. Таубер; Скотт Х. (1997). Генерация разнообразия: теория клонального отбора и рост молекулярной иммунологии (1-е изд. В мягкой обложке). Кембридж, Массачусетс: Harvard Univ. Нажмите. ISBN 0-674-00182-6.
- Авторы «Биология в контексте - спектр жизни», Питер Обюссон, Эйлин Кеннеди.
- Форсдайк Д. (1995). "Истоки теории клонального отбора иммунитета". Журнал FASEB. 9 (2): 164–66. Дои:10.1096 / fasebj.9.2.7781918. PMID 7781918. S2CID 38467403.
внешняя ссылка
- Анимация клонального отбора от института Уолтера и Элизы Холл.