Холобионт - Holobiont

А холобионт это сборка хозяин и многие другие виды, живущие в нем или вокруг него, вместе образующие отдельные экологическая единица.[1] Компоненты холобионта - это отдельные виды или бионты, а комбинированный геном всех бионтов это гологеном. Первоначально понятие холобионта было определено Доктор Линн Маргулис в ее книге 1991 года Симбиоз как источник эволюционных инноваций,[1] хотя концепция впоследствии эволюционировала с момента первоначального определения.[2] К голобионтам относятся хозяин, виром, микробиом, и другие члены, все из которых тем или иным образом способствуют функции целого.[3][4] К хорошо изученным холобионтам относятся: рифообразующие кораллы и люди.[5][6]
Существуют разногласия по поводу того, можно ли рассматривать холобионтов как единые эволюционные единицы.
Обзор
Холобионт - это совокупность видов, которые тесно связаны и имеют сложные взаимодействия, например, вид растения и его представители. микробиом.[1][7] Каждый вид, присутствующий в холобионте, является бионтом, и геномы всех бионтов, взятые вместе, представляют собой хологеном или «всеобъемлющую генную систему» холобионта.[8] Холобионт обычно включает эукариот хозяин и все симбиотический вирусы, бактерии, грибы и т. д., которые живут на нем или внутри него.[7]
Голобионты отличаются от суперорганизмы; суперорганизмы состоят из множества особей, иногда одного и того же вида, и этот термин обычно применяется к эусоциальный насекомые.[9][10] An колония муравьев может быть описан как суперорганизм, тогда как отдельный муравей и связанные с ним бактерии, грибы и т. д. являются холобионтом.[8] Нет сомнений в том, что симбиотические микроорганизмы имеют решающее значение для биологии и экологии хозяина, обеспечивая витамины, энергию и неорганические или органические питательные вещества, участвуя в защитных механизмах или управляя эволюцией хозяина.[11][12] Эти термины по-прежнему вызывают разногласия, и в некоторых публикациях они используются как синонимы.[6]
Компоненты холобионта
Хозяин: Хозяин холобионта обычно является многоклеточным. эукариот, например, растение или человек.[8] Известные хозяева, которые хорошо изучены, включают людей,[13] кораллы[5] и сосны.[14]
Микробиом: микробиом включает бактерии,[3] археи,[15] микроскопические грибы,[7] и микроскопические протисты.[3]
Виром: Все вирусы, включенные в холобионт, вместе именуются виром[16]
Грибы: в холобионты могут входить многоклеточные грибы, например арбускулярные микоризные грибы (AMF) в корнях растений.[7][4]
Фенотип холобионта

Холобионты - это существа, состоящие из хозяина и всех его симбиотических микробов.[2]
На диаграмме симбиотические микробы, поражающие холобионт. фенотип и вместе с хозяином окрашены в синий цвет, тогда как те, которые влияют на фенотип холобионта, но не эволюционировали совместно с хозяином, окрашены в красный цвет. Те, которые вообще не влияют на фенотип холобионта, окрашены в серый цвет. Микробы могут передаваться вертикально или горизонтально, могут быть получены из окружающей среды и могут быть постоянными или непостоянными в организме хозяина.[2]
Отсюда следует, что фенотипы холобионта могут изменяться во времени и пространстве, когда микробы входят в холобионт и выходят из него. Микробы в окружающей среде не являются частью холобионта (белого цвета). Затем гологеномы охватывают геномы хозяина и всех его микробов в любой заданный момент времени, причем отдельные геномы и гены попадают в одни и те же три функциональные категории: синий, красный и серый. Голобионты и гологеномы суть сущности, тогда как коэволюция или эволюция взаимодействий хозяин-симбионт - это процессы.[2]
Пример холобионтов
Люди и осы


Растения
Хотя большая часть работ по взаимодействию хозяев и микробов была сосредоточена на системах животных, таких как кораллы, губки или люди, существует значительный объем литературы по холобионтам растений.[20] Связанные с растениями микробные сообщества влияют на оба ключевых компонента приспособленности растений, роста и выживания,[4] и формируются доступностью питательных веществ и защитными механизмами растений.[7] Было описано несколько мест обитания микробов, связанных с растениями, включая ризоплан (поверхность корневой ткани), ризосфера (периферия корней), эндосфера (внутри растительной ткани) и филлосфера (общая площадь надземной поверхности).[12]
Растительный холобионт относительно хорошо изучен, с особым вниманием к сельскохозяйственным видам, таким как бобовые и зерна. Бактерии, грибы, археи, протисты и вирусы являются членами холобионта растения.[3]
Типы бактерий, которые, как известно, являются частью холобионта растений: Актинобактерии, Bacteroidetes, Фирмикуты, и Протеобактерии.[3] Например, азотфиксаторы такие как Азотобактер (Протеобактерии) и Бациллы (Firmicutes) значительно улучшают производительность растений.[3]
Грибы типа Аскомикота, Базидиомицеты, и Гломеромикота колонизируют ткани растений и обеспечивают множество функций для растения-хозяина.[3] Например, арбускулярные микоризные грибы (Glomeromycota) распространены среди групп растений и обеспечивают улучшенное усвоение питательных веществ, устойчивость к температуре и засухе, а также уменьшают возбудитель нагрузка.[21] Эпихлоя виды (Ascomycota) входят в состав овсяница луговая холобионт и обеспечивают устойчивость травоядных, производя алкалоиды спорыньи, что вызывает эрготизм у млекопитающих.[22]
Протистские представители холобионта растения менее изучены, при этом большая часть знаний ориентирована на патогены. Однако есть примеры комменсалистический ассоциации растений-протистов, такие как Фитомонады (Trypanosomatidae ).[23]
морской
Кораллы-строители рифов - это холобионты, в том числе и сам коралл (эукариотический беспозвоночный в классе Антозоа ), фотосинтетический динофлагелляты называется зооксантеллы (Симбиодиниум ), а также связанных бактерий и вирусов.[5] Модели коэволюции существуют для сообществ коралловых микробов и филогении кораллов.[24]
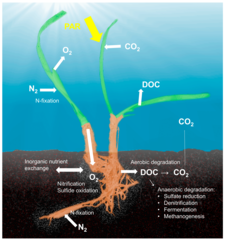

Воздействие стрессоров

(1) физиология и иммунитет хозяина
(2) состав и плотность симбиотического сообщества. Стрессоры также могут косвенно влиять на симбиотическое сообщество, изменяя физиологию хозяина (которая представляет собой симбиотическую нишу) и иммунное состояние хозяина. И наоборот, симбионты могут буферизовать стрессоры за счет обеспечения питательными веществами, физиологической толерантности и защиты от естественных врагов хозяина.[28]

(1) характеристики фактора стресса, такие как его частота или амплитуда, а также его сочетание с другим фактором стресса, которое может привести к аддитивному, синергетическому или антагонистическому взаимодействию
(2) способ передачи симбионтов
(3) специфика и эффективность данного механизма буферизации, а также чистый баланс между его стоимостью и его выгодой.[28]
Полемика
В последние годы были разработаны мощные, но относительно недорогие инструменты для характеристики микробных сообществ, в том числе высокопроизводительное секвенирование такие технологии как полногеномное секвенирование. Эти технологические достижения привели к взрыву интереса к микробной экологии и эволюции взаимоотношений между микробами и хозяевами. Некоторые исследователи задаются вопросом, нужна ли концепция холобионта и соответствует ли она сложностям отношений между хозяином и симбионтом.[29] В 2016 году Дуглас и Веррен оспорили концепцию, согласно которой «холобионт (хозяин плюс его микробиом) и составляющий его хологеном (совокупность геномов в холобионте) являются единицей отбора, и поэтому эта единица имеет свойства, аналогичные свойствам отдельного организма. ".[30] Они утверждают, что «концепция хологенома бесполезна для изучения взаимодействий хозяина с резидентными микроорганизмами, потому что она сосредоточена на одном уровне отбора (холобионт) и, как следствие, связана с кооперативными и интегративными функциями систем хозяин-микроб для исключение других видов взаимодействий, включая антагонизм между микроорганизмами и конфликты между хозяином и микробными партнерами ».[30]
Концепция холобионта и, как следствие, концепция гологенома остаются противоречивыми, особенно в отношении хозяина и его микробиома как единой эволюционной единицы.[31] Чтобы подтвердить концепцию холобионта с эволюционной точки зрения, необходимы новые теоретические подходы, которые признают различные уровни, на которых естественный отбор может действовать в контексте взаимодействий микробиом-хозяин. Например, отбор может происходить на уровне холобионта, когда может поддерживаться трансгенерационная ассоциация между конкретными генотипами хозяина и симбионта.[31]
Тем не менее, концепция холобионта привела к смещению акцента на симбиозах с участием одного микробного партнера и одного хозяина (кальмаров и люминесцентных Аливибрио, бобовые и Ризобий, тля и Бухнера ) к большему интересу к симбиозам в сложных многопартнерских консорциумах (кишечные системы животных, морские беспозвоночные, эпифиты растений и водорослей, микробно-микробные взаимодействия в почве, водные биомы).[31] Более того, есть осознание того, что даже относительно хорошо изученные бинарные симбиозы, такие как тля и Buchnera, являются более сложными с рядом различных факультативных симбионтов, способствующих устойчивости к паразитам,[32] расширение использования принимающих растений[33] и температурная адаптация.[34][31]
Смотрите также
использованная литература
- ^ а б c Маргулис, Линн; Фестер, Рене (1991). Симбиоз как источник эволюционных инноваций. MIT Press. ISBN 9780262132695.
- ^ а б c d е Тайс, К.Р., Дайли, Н.М., Классен, Дж. Л., Брукер, Р. М., Бейнс, Дж. Ф., Бош, Т. К., Крайан, Дж. Ф., Гилберт, С. Ф., Спокойной ночи, К. Дж., Ллойд, Э. и Сапп, Дж. (2016) «Правильное понимание концепции гологенома: экоэволюционная структура для хозяев и их микробиомов». Msystems, 1(2). Дои:10.1128 / м Системы.00028-16.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ а б c d е ж г Булгарелли, Д., Шлаэппи, К., Спапен, С., Ван Темат, Э.В.Л. и Schulze-Lefert, P. (2013) "Структура и функции бактериальной микробиоты растений". Ежегодный обзор биологии растений, 64: 807–838. Дои:10.1146 / annurev-arplant-050312-120106.
- ^ а б c Ванденкоорнхейз, Филипп; Квайзер, Ахим; Дюамель, Мари; Ле Ван, Амандин; Дюфрен, Алексис (01.06.2015). «Важность микробиома холобионта растения». Новый Фитолог. 206 (4): 1196–1206. Дои:10.1111 / nph.13312. ISSN 1469-8137. PMID 25655016.
- ^ а б c Ноултон, Н. и Ровер, Ф. (2003) «Многовидовые микробные мутуализмы на коралловых рифах: хозяин как среда обитания». Американский натуралист, 162(S4): S51-S62. Дои:10.1086/378684.
- ^ а б Крамер, Питер; Брессан, Паола (2015). «Люди как суперорганизмы: как микробы, вирусы, импринтированные гены и другие эгоистичные сущности формируют наше поведение». Перспективы психологической науки. 10 (4): 464–481. Дои:10.1177/1745691615583131. ISSN 1745-6916. PMID 26177948. S2CID 12954636. Бесплатный, полный текст.
- ^ а б c d е Санчес-Каньисарес, К., Хоррин, Б., Пул, П.С. и Ткач А. (2017) «Понимание холобионта: взаимозависимость растений и их микробиома». Текущее мнение в микробиологии, 38: 188–196. Дои:10.1016 / j.mib.2017.07.001.
- ^ а б c Bordenstein, Seth R .; Тайс, Кевин Р. (2015). «Биология хозяина в свете микробиома: десять принципов голобионтов и гологеномов». ПЛОС Биол. 13 (8): e1002226. Дои:10.1371 / journal.pbio.1002226. ISSN 1545-7885. ЧВК 4540581. PMID 26284777.
- ^ Гордон, Дж., Ноултон, Н., Релман, Д.А., Ровер, Ф. и Юл, М. (2013) "Суперорганизмы и холобионты". Микроб, 8(4): 152–153. Дои:10.1128 / microbe.8.152.1.
- ^ Уиллер, WM (1928). Социальные насекомые, их происхождение и эволюция. Харкорт Брейс.
- ^ Розенберг, Э. и Зильбер-Розенберг, И. (2016) "Микробы управляют эволюцией животных и растений: концепция гологенома". MBio, 7(2). Дои:10,1128 / мБио.01395-15.
- ^ а б c Угарелли К., Чакрабарти С., Лаас П. и Стингл У. (2017) «Холобионт из морских водорослей и его микробиом». Микроорганизмы, 5(4): 81. Дои:10.3390 / микроорганизмы 5040081. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Гухте, Маартен ван де, Эрве М. Блоттьер и Жоэль Доре. «Люди как холобионты: значение для профилактики и лечения». Микробиом 6, вып. 1 (декабрь 2018 г.): 81. https://doi.org/10.1186/s40168-018-0466-8.
- ^ Хаккард, Стефан и Кристофер В. Шадт (2015) «На пути к целостному пониманию полезных взаимодействий через Populus Микробиом ". Новый Фитолог, 205(4): 1424–30. Дои:10.1111 / nph.13133.
- ^ Хассани, М.А., Дуран, П. и Хаккард, С. (2018) "Микробные взаимодействия внутри холобионта растения". Микробиом, 6(1): 58. Дои:10.1186 / s40168-018-0445-0
- ^ Grasis, J.A. (2017) «Взаимозависимость вирусов и холобионта». Границы иммунологии, 8: 1501. Дои:10.3389 / fimmu.2017.01501.
- ^ Постлер, Т. и Гош, С. (2017) «Понимание холобионта: как микробные метаболиты влияют на здоровье человека и формируют иммунную систему». Клеточный метаболизм, 26(1): 110–130. Дои:10.1016 / j.cmet.2017.05.008.
- ^ Ричардсон, Л.А. (2017) "Эволюция как холобионт". PLoS биология, 15(2): e2002168. Дои:10.1371 / journal.pbio.2000225. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Брукс, А.В., Коль, К.Д., Брукер, Р.М., ван Опсталь, Э.Дж. и Борденштейн, С. (2016) «Филосимбиоз: взаимосвязи и функциональные эффекты микробных сообществ на протяжении истории эволюции хозяина». PLoS биология, 14(11): e2000225. Дои:10.1371 / journal.pbio.2000225.
- ^ Зильбер-Розенберг, И. и Розенберг, Э. (2008) "Роль микроорганизмов в эволюции животных и растений: теория эволюции гологенома". Обзор микробиологии FEMS, 32(5): 723–735. Дои:10.1111 / j.1574-6976.2008.00123.x.
- ^ Бегум, Н., Цин, К., Ахангер, М.А., Раза, С., Хан, М.И., Ахмед, Н., Ашраф, М., и Чжан, Л. (2019) «Роль арбускулярных микоризных грибов в регуляции роста растений. : Влияние на устойчивость к абиотическому стрессу ». Границы науки о растениях, 10: 1068. Дои:10.3389 / fpls.2019.01068.
- ^ Герре, П. (2015) «Алкалоиды спорыньи, продуцируемые эндофитными грибами этого рода. Эпихлоя". Токсины, 7(3): 773–790. Дои:10.3390 / токсины7030773.
- ^ Швельм, А., Бадстебер, Дж., Булман, С., Дезуань, Н., Этемади, М., Фаллон, Р. Э., Гачон, К. М., Легрев, А., Лукеш, Дж., Мерц, У. и Ненарокова, А., 2018. Не в твоей обычной десятке: протисты, поражающие растения и водоросли. Молекулярная патология растений, 19 (4), стр. 1029-1044. Дои:10.1111 / mpp.12580.
- ^ Поллок, Ф. Джозеф; МакМиндс, Райан; Смит, стили; Борн, Дэвид Дж .; Уиллис, Бетт Л .; Медина, Моника; Тербер, Ребекка Вега; Заневельд, Джесси Р. (22.11.2018). «Связанные с кораллами бактерии демонстрируют филосимбиоз и кофилогению». Nature Communications. 9 (1): 4921. Дои:10.1038 / с41467-018-07275-х. ISSN 2041-1723. ЧВК 6250698. PMID 30467310.
- ^ Томпсон, Дж. Р., Ривера, Х. Э., Клоек, С. Дж. И Медина, М. (2015) «Микробы в коралловом холобионте: партнеры в процессе эволюции, развития и экологических взаимодействий». Границы клеточной и инфекционной микробиологии, 4: 176. Дои:10.3389 / fcimb.2014.00176.
- ^ Пита, Л., Рикс, Л., Слаби, Б.М., Франке, А., Хентшель, У. (2018) «Губчатый холобионт в меняющемся океане: от микробов к экосистемам». Микробиом, 6(1): 46. Дои:10.1186 / s40168-018-0428-1. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Кавальканти, Г.С., Шукла, П., Моррис, М., Рибейро, Б., Фоли, М., Доан, М.П., Томпсон, К.С., Эдвардс, М.С., Динсдейл, Е.А. и Томпсон, Ф. (2018) «Холобионты родолитов в меняющемся океане: взаимодействия хозяина и микробов опосредуют устойчивость коралловых водорослей при закислении океана». BMC Genomics, 19(1): 1–13. Дои:10.1186 / s12864-018-5064-4. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ а б Бенар, А., Вавр, Ф. и Кремер, Н. (2020) «Стресс и симбиоз: голова или решка?». Границы экологии и эволюции, 8: 167. Дои:10.3389 / fevo.2020.00167. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Moran, Nancy A .; Слоан, Дэниел Б. (2015). «Концепция гологенома: полезная или пустая?». PLOS Биология. 13 (12): e1002311. Дои:10.1371 / journal.pbio.1002311. ISSN 1545-7885. ЧВК 4670207. PMID 26636661.
- ^ а б Дуглас, А.Е., Веррен, Дж. Х. (2016) «Дыры в хологеноме: почему симбиозы хозяин-микроб не являются холобионтами». MBio, 7(2). Дои:10.1128 / mBio.02099-15.
- ^ а б c d Иган, С., Фукацу, Т. и Францино, М.П. (2020) «Возможности и проблемы исследования микробного симбиоза в эру микробиома». Границы микробиологии, 11. Дои:10.3389 / fmicb.2020.01150. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Оливер К. М., Рассел Дж. А., Моран Н. А. и Хантер М. С. (2003) "Факультативные бактериальные симбионты у тлей придают устойчивость к паразитическим осам". Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ., 100: 1803–1807. Дои:10.1073 / пнас.0335320100.
- ^ Цучида Т., Кога Р. и Фукацу Т. (2004) "Специализация растения-хозяина определяется факультативным симбионтом". Наука, 303: 1989. Дои:10.1126 / science.1094611.
- ^ Montllor, C.B., Maxmen, A., и Purcell, A.H. (2002) «Факультативные бактериальные эндосимбионты приносят пользу гороховой тле. Acyrthosiphon pisum при тепловом стрессе ». Ecol. Энтомол., 27: 189–195. Дои:10.1046 / j.1365-2311.2002.00393.x.
| Группы |  | |
|---|---|---|
| Микробиология | ||
| Экология | ||
| Растения | ||
| морской | ||
| Связанные с людьми | ||
| Методы | ||
| Другой | ||
| ||