Глубокая биосфера - Deep biosphere
В глубокая биосфера является частью биосфера который находится ниже первых нескольких метров поверхности. Он простирается как минимум на 5 км ниже поверхности материка и на 10,5 км ниже поверхности моря при температурах, которые могут превышать 100 ° C. Включает в себя все три области жизни и генетическое разнообразие соперников на поверхности.
Первые признаки глубинной жизни были получены в результате исследований нефтяных месторождений в 1920-х годах, но не было уверенности в том, что эти организмы были местными, пока в 1980-х годах не были разработаны методы предотвращения загрязнения с поверхности. Сейчас пробы собираются в глубоких шахтах и научное бурение программы в океане и на суше. Были созданы глубокие обсерватории для более обширных исследований.
У поверхности живые организмы потребляют органическое вещество и дышат кислородом. Ниже они недоступны, поэтому в них используются «съедобные» (доноры электронов ) Такие как водород (высвобождается из горных пород в результате различных химических процессов), метан (CH4), уменьшенный сера соединения и аммоний (NH4). Они «дышат» акцепторы электронов Такие как нитраты и нитриты, марганец и оксиды железа, окисленные соединения серы и углекислый газ (CO2). На больших глубинах очень мало энергии, поэтому метаболизм в миллион раз медленнее, чем на поверхности. Клетки могут жить тысячи лет до деления, и их возраст неизвестен.
Недра составляет около 90% биомасса в двух сферах жизни, Археи и Бактерии, и 15% от общего количества для биосферы. Встречаются также эукарии, в том числе некоторая многоклеточная жизнь (нематоды, грибы, плоские черви, коловратки, кольчатые червя и членистоногие ). Также присутствуют вирусы, которые заражают микробы.
Определение
Глубокая биосфера - это экосистема организмов и их жизненное пространство в глубоких недрах.[1] Для морского дна рабочее определение глубокие недра область, не биотурбированная животными; это обычно примерно на метр или более ниже поверхности.[2] На континентах он ниже нескольких метров, не считая почвы.[3] Организмы в этой зоне иногда называют внутриземные.[4][5]
История
На Чикагский университет В 1920-х годах геолог Эдсон Бастин заручился помощью микробиолога Фрэнка Грира, пытаясь объяснить, почему вода, добытая на нефтяных месторождениях, содержит сероводород и бикарбонаты. Эти химические вещества обычно создаются бактериями, но вода поступала из глубины, где температура и давление считались слишком высокими для поддержания жизни. Они смогли культивировать анаэробные сульфатредуцирующие бактерии из воды, что свидетельствует о бактериальном происхождении химикатов.[6][7][8]
Также в 1920-х гг. Чарльз Липман, микробиолог в Калифорнийский университет в Беркли, заметили, что бактерии, которые были запечатаны в бутылках в течение 40 лет, можно реанимировать - явление, теперь известное как ангидробиоз. Он задавался вопросом, верно ли то же самое в отношении бактерий в угольных пластах. Он стерилизовал образцы угля, смачивал их, измельчал, а затем преуспел в выращивании бактерий из угольной пыли. Одна процедура стерилизации, обжигание угля при 160 градусах Цельсия в течение 50 часов, фактически стимулировала их рост. Он опубликовал результаты в 1931 году.[4][8]
Первые исследования подземной жизни были проведены Клод Э. Зобелл, «отец морской микробиологии»,[9] в конце 1930-х - 1950-х гг. Хотя глубина отбора керна была ограничена, микробы были обнаружены везде, где отбирались образцы отложений.[10][11] С увеличением глубины аэробы уступил место анаэробы.[12]

Большинство биологов отклонили подповерхностные микробы как загрязнение, особенно после погружения в воду. Элвин затонул в 1968 году, и ученые сбежали, оставив свой обед позади. Когда Элвин был восстановлен, обеды не показали никаких признаков микробного разложения.[9] Это усиливало представление о глубоком море (и, соответственно, о недрах) как о безжизненной пустыне. Изучение глубинной биосферы оставалось бездействующим на протяжении десятилетий, за исключением некоторых советских микробиологов, которые начали называть себя геомикробиологи.[8]
Интерес к подземной жизни возобновился, когда Министерство энергетики США искал безопасный способ захоронения ядерных отходов, и Фрэнк Дж. Воббер поняли, что микробы под поверхностью могут либо помочь, разлагая захороненные отходы, либо мешать, пробивая герметичные контейнеры. Он создал Программу изучения недр для изучения глубинной жизни. Для решения проблемы загрязнения было разработано специальное оборудование, минимизирующее контакт между образцом керна и буровой раствор что смазывает сверло. Кроме того, в жидкость были добавлены индикаторы, чтобы указать, проникла ли она в керн. В 1987 году несколько скважины были пробурены возле Сайт реки Саванна, и было обнаружено, что микроорганизмы многочисленны и разнообразны, по крайней мере, на 500 метров ниже поверхности.[11]
С 1983 по 2003 год микробиологи проанализировали численность клеток в кернах буровых скважин. Программа морского бурения.[10] Группа во главе с Джоном Парксом из Бристольский университет сообщенные концентрации 104 до 108 клеток на грамм осадка на глубине до 500 метров (в сельскохозяйственных почвах содержится около 109 клеток на грамм).[13] Первоначально это было встречено скептически, и им потребовалось четыре года, чтобы опубликовать свои результаты.[9] В 1998 году Уильям Уитман и его коллеги опубликовали сводку данных за двенадцать лет в Труды Национальной академии наук.[13] По их оценкам, до 95% всех прокариоты (археи и бактерии) живут в глубоких недрах, из них 55% - в морских недрах и 39% - в земных недрах.[10] В 2002 году ODP Leg 201 был первым, кто был мотивирован поиском глубокой жизни. Большая часть предыдущей разведки проводилась на континентальных окраинах, поэтому целью было бурение в открытом океане для сравнения.[14][4]
Научные методы
Современное понимание подповерхностной биологии стало возможным благодаря многочисленным достижениям в технологии сбора образцов, полевого анализа, молекулярной науки, культивирования, визуализации и вычислений.[12]
Сбор образцов


Дно океана отбирается путем бурения скважин и сбора керна. Методы должны быть адаптированы к различным виды рока, а стоимость сверления ограничивает количество отверстий, которые можно просверлить.[16] Микробиологи использовали научные программы бурения: Программа морского бурения (ODP), который использовал ДЖОИДЕС Разрешение буровая платформа и Комплексная программа морского бурения (IODP), который использовал японский корабль Тикю.[12]
Например, глубокие подземные шахты Золотые рудники Южной Африки и Медно-цинковый рудник Пюхясалми в Финляндия, предоставили возможность исследовать глубинную биосферу.[17][18] Образцы глубинных недр также отбирались на выбранных или предполагаемых площадках для захоронения ядерных отходов (например, Юкка Маунтин и Опытная установка по изоляции отходов В Соединенных Штатах, Äspö и прилегающих районах Швеции, Онкало и прилегающие районы в Финляндии, Mont Terri в Швейцарии).[12] Научному бурению континентальных глубоких недр способствует Международная континентальная программа научного бурения (ICDP).[19]
Чтобы обеспечить непрерывный подземный отбор проб, были созданы различные обсерватории. На дне океана комплект для модернизации Circulation Obviation Retrofit Kit (CORK) закрывает скважину, чтобы перекрыть приток морской воды.[20] Усовершенствованная версия CORK может изолировать несколько секций буровой скважины с помощью упаковщики, резиновые трубки, которые надуваются, чтобы запечатать пространство между бурильной колонны и стенка скважины.[21][22] В отложениях простой тросовый прибор для измерения параметров на месте (SCIMPI) предназначен для того, чтобы оставаться и проводить измерения после обрушения ствола скважины.[23]
Пакеры также используются в континентальных недрах,[24] вместе с такими устройствами, как проточные на месте реактор (FTISR). Для отбора жидкостей с этих участков используются различные методы, включая пассивные газовые пробоотборники, системы U-образных трубок и осмотический пробоотборники газа.[12] В узких (менее 50 миллиметров) отверстиях, полиамид трубки с обратным клапаном можно опускать для отбора проб всего столба жидкости.[25][26]
Полевой анализ и манипуляции
Некоторые методы анализа микробов на месте а не извлекать их. В биогеофизика, влияние микробов на свойства геологических материалов определяется дистанционно с помощью электрических сигналов. Микробы можно пометить с помощью стабильного изотопа, например углерод-13 а затем повторно закачивают в землю, чтобы посмотреть, куда они денутся.[12] Метод «двухтактный» включает закачку жидкости в водоносный горизонт и извлечение смеси закачиваемой жидкости с грунтовыми водами; последние затем можно проанализировать, чтобы определить, какие химические реакции произошли.[27]
Молекулярные методы и выращивание
Методы современной молекулярной биологии позволяют извлекать нуклеиновые кислоты, липиды и белки из клеток, Секвенирование ДНК, а также физико-химический анализ молекул с использованием масс-спектрометрии и проточной цитометрии. Используя эти методы, можно многое узнать о микробных сообществах, даже если их невозможно культивировать.[12] Например, на Richmond Mine в Калифорнии ученые использовали секвенирование дробовика чтобы идентифицировать четыре новых вида бактерий, три новых вида архей (известных как Архей Ричмонд Майн ацидофильные наноорганизмы ) и 572 белка, уникальных для бактерий.[28][29]
Геохимические методы
Глубинные микроорганизмы изменяют химию своего окружения. Они потребляют питательные вещества и отходы продуктов из метаболизм. Таким образом, мы можем оценить активность глубинных микроорганизмов, измерив химический состав образцов под полом. Дополнительные методы включают измерение изотоп состав химикатов или связанных минералы.[30][неудачная проверка ]
Условия для жизни
Чтобы жизнь имела метаболическую активность, она должна уметь пользоваться термодинамическое неравновесие в окружающей среде. Это может произойти, когда мантийные породы, богатые минералами оливин подвергаются воздействию морской воды и вступают в реакцию с ней с образованием серпентиновых минералов и магнетит.[31] Неравновесные условия также связаны с гидротермальные источники, вулканизм, и геотермальная активность. Другие процессы, которые могут обеспечить среду обитания для жизни, включают развитие фронта валков в рудных телах,[примечание 1] субдукция, клатрат метана образование и разложение, вечная мерзлота таяние инфракрасный радиационная и сейсмическая активность. Люди также создают новые среды обитания для жизни, особенно с помощью очистка от загрязняющих веществ в недрах.[10]
Источники энергии
Жизнь требует достаточно энергии, чтобы построить аденозинтрифосфат (АТФ). Там, где есть солнечный свет, основными процессами улавливания энергии являются фотосинтез (который использует энергию солнечного света преобразование углекислый газ в Органические молекулы ) и дыхание (который потребляет эти молекулы и выделяет углекислый газ). Под землей основным источником энергии является химическая редокс (восстановительно-окислительные) реакции. Это требует доноры электронов (соединения, которые могут окисляться) и акцепторы электронов (соединения, которые можно восстановить). Пример такой реакции - окисление метана:
- CH4 + O2 → CO2 + 2 часа2О
Здесь CH4 донор и O2 акцептор.[33] Доноров можно считать «съедобными», а получателей - «дышащими».[34]
Количество энергии, которое выделяется при метаболической реакции, зависит от окислительно-восстановительный потенциал химических веществ. Доноры электронов имеют отрицательный потенциал. От самого высокого до самого низкого окислительно-восстановительного потенциала, некоторые общие доноры, доступные в недрах, - это органические вещества, водород, метан, восстановленные соединения серы, восстановленные соединения железа и аммоний. От самых отрицательных до минимальных, некоторые акцепторы - кислород, нитраты и нитриты, оксиды марганца и железа, окисленные соединения серы и диоксид углерода. [33]
Из доноров электронов наиболее отрицательным окислительно-восстановительным потенциалом обладает органическое вещество. Он может состоять из отложений из регионов, где есть солнечный свет, или из месторождений. Свежий материал утилизировать легче, чем выдержанный. Земное органическое вещество (в основном из растений), как правило, труднее обрабатывать, чем морское (фитопланктон). Некоторые организмы расщепляют органические соединения, используя ферментация и гидролиз, давая возможность другим преобразовать его обратно в диоксид углерода. Водород - хороший источник энергии, но конкуренция делает его дефицитным. Он особенно богат гидротермальными жидкостями там, где его получают серпентинизацией. Несколько видов могут сочетать ферментацию с метаногенез и окисление железа с расходом водорода. Метан в основном содержится в морских отложениях, в газообразной форме (растворенной или свободной) или в гидраты метана. Около 20% поступает из абиотических источников (разложение органических веществ или серпентинизация) и 80% из биотических источников (которые снижают количество органических соединений, таких как диоксид углерода, оксид углерода и др.). ацетат ). Более 90% метана окисляется микробами до того, как достигнет поверхности; эта деятельность является «одним из наиболее важных средств контроля выбросов парниковых газов и климата на Земле».[33] Восстановленные соединения серы, такие как элементарная сера, сероводород (ЧАС2Песок пирит (FeS2) находятся в гидротермальные источники в базальтовой коре, где они выпадают в осадок, когда богатые металлами флюиды контактируют с морской водой. Восстановленные соединения железа в отложениях в основном осаждаются или образуются анаэробным восстановлением оксиды железа.[33]
Акцептором электронов с наивысшим окислительно-восстановительным потенциалом является кислород. Произведенный фотосинтезом, он переносится на дно океана. Там он быстро принимается, если органического материала много, и может присутствовать только в верхних нескольких сантиметрах. В отложениях с низким содержанием органических веществ его можно найти на больших глубинах, вплоть до океанической коры. Нитраты могут образовываться в результате разложения органических веществ или фиксации азота.[33] Кислород и нитраты получаются в результате фотосинтеза, поэтому подземные сообщества, которые их используют, не являются полностью независимыми от поверхности.[35]
Питательные вещества
Вся жизнь требует углерода, азота, фосфора и некоторых микроэлементов, таких как никель, молибден и ванадий. Более 99,9% углерода Земли хранится в коре и вышележащих отложениях, но доступность этого углерода может зависеть от степени окисления окружающей среды. Органический углерод, азот и фосфор в основном содержатся в земных отложениях, которые накапливаются в основном на окраинах континентов. Органический углерод в основном вырабатывается на поверхности океанов с фотосинтез или смывается в океаны с земными отложениями. Лишь небольшая часть производится в глубоких морях с хемосинтез. Когда органический углерод опускается с поверхности океана на морское дно, большая часть органического углерода потребляется организмами в морской воде. Лишь небольшая часть этого тонущего органического углерода может достигнуть морского дна и быть доступной для глубинной биосферы.[30][неудачная проверка ] Глубже в морских отложениях содержание органических веществ падает.[30] Фосфор поглощается оксигидроксиды железа когда базальты и сульфидные породы выветриваются, что ограничивает его доступность.[36] Доступность питательных веществ ограничивает глубинную биосферу, определяя, где и какой тип глубинных организмов могут процветать.
Давление

Атмосферное давление 101килопаскали (кПа). В океане давление увеличивается со скоростью 10,5 кПа на метр глубины, поэтому на типичной глубине морского дна (3800 метров) давление составляет 38 мегапаскалей (МПа). На этих глубинах температура кипения воды превышает 400 ° C. На дне Марианской впадины давление 110 МПа. В литосфере давление увеличивается на 22,6 кПа на метр.[36][37] Глубокая биосфера выдерживает давление, намного превышающее давление на поверхности Земли.[30]
Повышенное давление сжимает липиды, делая мембраны менее жидкими. В большинстве химических реакций продукты занимают больше объема, чем реагенты, поэтому реакции тормозятся давлением.[37] Тем не менее, некоторые исследования утверждают, что клетки с поверхности все еще активны при давлении в 1 гигапаскаль (ГПа). Это также пьезофилы для которых оптимальный рост происходит при давлениях более 100 МПа,[36] а некоторые не растут при давлении ниже 50 МПа.[37]
По состоянию на 2019 год большая часть образцов организмов, отобранных из глубин океана и недр, подвергается декомпрессии, когда их выносят на поверхность. Это может повредить клетки разными способами, а эксперименты с поверхностным давлением дают неточную картину микробной активности в глубокой биосфере.[38][39][40] Подводный манипулятор пробоотборника под давлением (PUSH50) был разработан для обслуживания на месте давление во время отбора проб и после этого в лаборатории.[41]
Температура
Высокие температуры вызывают стресс у организмов, увеличивая скорость процессов, которые повреждают важные молекулы, такие как ДНК и аминокислоты. Это также увеличивает потребность в энергии для восстановления этих молекул.[42] Однако клетки могут реагировать, изменяя структуру этих молекул, чтобы стабилизировать их.[36][37][43]
Микробы могут выжить при температуре выше 100 ° C, если давление достаточно высокое, чтобы вода не закипела. Самая высокая температура, при которой организм был выращен в лаборатории, составляет 122 ° C,[33][36] под давлением 20 и 40 мегапаскалей.[44] Теоретические оценки максимальной температуры, которая может поддерживать жизнь, составляют около 150 ° C.[45] Изотерма 120 ° C может быть менее 10 метров в глубине срединно-океанических хребтов и подводных гор, но в других средах, например в глубоководных желобах, она может достигать нескольких километров.[36] Около 39% объема океанических отложений находится при температуре от 40 ° C до 120 ° C.[45]
Рекордный термофил, Methanopyrus kandlerii, был изолирован от гидротермального источника.[45] Гидротермальные источники дают много энергии и питательных веществ. Несколько групп архей и бактерий процветают на мелководье морского дна при температуре от 80 до 105 ° C. В средах с более ограниченным энергопотреблением ожидается, что ячейки будут иметь более низкие пределы температуры. Хотя микробные признаки были обнаружены при температурах до 100 ° C в керосиновых отложениях,[46] попытки изолировать организмы потерпели неудачу. В глубоких нефтяных коллекторах с температурой выше 80 ° C микробная активность не наблюдалась, а предел может составлять всего 60 ° C в глубоко захороненных отложениях.[42]
Жизнь с ограничением энергии
В большей части недр организмы живут в условиях крайней нехватки энергии и питательных веществ.[30] Это далеко от условий, в которых клетки культивируются в лабораториях. Лабораторная культура проходит ряд предсказуемых фаз. После небольшой фазы задержки наступает период экспоненциального роста, в течение которого популяция может удвоиться всего за 20 минут. Далее следует фаза смерти, в которой почти все клетки отмирают. Остальные входят в длительную стационарную фазу, в которой они могут существовать годами без дополнительного добавления субстрата. Тем не менее, каждая живая клетка имеет от 100 до 1000 мертвых клеток, которыми можно питаться, поэтому у них все еще есть много питательных веществ по сравнению с подповерхностными слоями.[1]
В недрах клетки катаболизировать (расщепляют молекулы для получения энергии или строительных материалов) в 10 000–1 000 раз медленнее, чем на поверхности. Для биомассы могут потребоваться столетия или тысячелетия. перевернуть. Не существует известного предела возрасту, которого могут достичь клетки. Присутствующие вирусы могут убивать клетки, и эукариоты могут пастись на них, но свидетельств этого нет.[1]
Трудно установить четкие пределы энергии, необходимой для поддержания жизни клеток, но не для их роста.[30] Им нужна энергия для выполнения определенных основных функций, таких как поддержание осмотического давления и поддержание макромолекул, таких как ферменты и РНК (например., корректура и синтез). Однако лабораторные оценки необходимой энергии на несколько порядков больше, чем запас энергии, который, по-видимому, поддерживает жизнь под землей.[1]
Сначала считалось, что большинство подземных ячеек бездействующий. Однако для выхода из состояния покоя требуется дополнительная энергия. Это не лучшая стратегия в среде, где источники энергии стабильны на протяжении миллионов лет, но медленно сокращаются. Имеющиеся данные свидетельствуют о том, что большинство подземных ячеек активны и жизнеспособны.[1]
Низкоэнергетическая среда способствует минимальному саморегулированию клеток, поскольку в окружающей среде нет изменений, на которые они должны реагировать. Могут быть специалисты с низким энергопотреблением. Однако сильных вряд ли будет эволюционное давление чтобы такие организмы могли развиваться из-за низкой текучести и потому, что окружающая среда является тупиком.[1]
Разнообразие
В биомасса в глубоких недрах составляет около 15% от общего количества биосферы.[3] Жизнь от всех трех области жизни (Археи, Бактерии, и Эукария ) были обнаружены в глубоких недрах;[47] действительно, глубокие недра составляют около 90% всей биомассы архей и бактерий.[3] Генетическое разнообразие по крайней мере так же велико, как на поверхности.[47]
В океане виды планктона распространены по всему миру и постоянно откладываются почти повсеместно. Совершенно разные сообщества встречаются даже на дне океана, и разнообразие видов уменьшается с глубиной.[48] Тем не менее, некоторые таксоны все еще широко распространены в недрах.[49] В морских отложениях основные бактериальный тип находятся "Candidatus Атрибактерии »(ранее OP9[50] и СП1[51]),[52] Протеобактерии, Хлорофлекси и Планктомицеты.[49] Члены архей были впервые идентифицированы с помощью метагеномный анализ; некоторые из них с тех пор были выращены и получили новые имена. Группа глубоководных архей (DSAG) стала морской бентосной группой B (MBG-B) и теперь является предлагаемым типом "Lokiarchaeota ".[48] Наряду с бывшей группой древних архей (AAG) и группой морских гидротермальных источников (MHVG), «Lokiarchaeota» является частью кандидата в суперфилум, Асгард.[53] Другие типы "Батиархей "(ранее - Разная кренархеотическая группа), Таумархеота (ранее морская группа I),[48] и Euryarchaeota (включая "Гадесархея ", Археоглобалы и Thermococcales ).[45] Также представлена родственная клада, анаэробные метанотрофные археи (ANME).[29] Другие бактериальные типы включают: Термотоги.[45]
В континентальных недрах основными бактериальными группами являются Proteobacteria и Фирмикуты в то время как археи в основном Метаномикробия и Thaumarchaeota.[54] К другим типам относятся «Bathyarchaeota» и «Aigarchaeota ", в то время как бактериальные типы включают Водные и Нитроспиры.[45]
Эукарии в глубокой биосфере включают в себя некоторую многоклеточную жизнь. В 2009 г. вид нематода, Halicephalobus mephisto, был обнаружен в трещинах горных пород более чем в километре от южноафриканского золотого рудника. По прозвищу «дьявольский червь»,[55] возможно, он был вытеснен землетрясениями вместе с поровой водой.[56] С тех пор были обнаружены и другие многоклеточные организмы, в том числе грибы, Платигельминты (плоские черви), Коловратка, Аннелида (кольчатые черви) и Членистоногие.[57][58][59][60][61][62] Однако их диапазон может быть ограничен, поскольку стеролы, необходимые для построения мембран у эукарий, нелегко создать в анаэробных условиях.[12]
Вирусы также присутствуют в больших количествах и заражают широкий спектр микробов в глубокой биосфере. Они могут вносить значительный вклад в оборот клеток и передачу генетической информации между клетками.[12]
Среды обитания
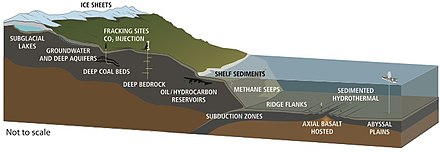
Жизнь была обнаружена на глубинах 5 км на континентах и 10,5 км под поверхностью океана.[47] В 1992 г. Томас Голд подсчитали, что если бы предполагаемое поровое пространство земного массива на глубине до 5 км было заполнено водой, и если бы 1% этого объема составляла микробная биомасса, было бы достаточно живого вещества, чтобы покрыть поверхность суши слоем толщиной 1,5 м. .[63] Предполагаемый объем глубинной биосферы составляет 2–2,3 миллиарда кубических километров, что примерно вдвое превышает объем океанов.[64]
дно океана
Основными типами местообитаний ниже морского дна являются отложения и вулканическая порода. Последний может быть частично изменен и сосуществовать с продуктами его изменения, такими как сульфиды и карбонаты. В горных породах химические вещества в основном переносятся через водоносный горизонт система, которая циклирует всю воду океана каждые 200000 лет. В отложениях ниже верхних нескольких сантиметров химические вещества распространяются в основном гораздо более медленным процессом. распространение.[36]
Отложения
Почти все морское дно покрыто морскими отложениями. Их толщина может варьироваться от сантиметров у хребтов океана до глубины более 10 километров. траншеи. Посреди океана, кокколиты и снаряды, располагающиеся с поверхности, образуют сочится, а прибрежные отложения переносятся реками с континентов. Также вносят свой вклад минералы из гидротермальных источников и переносимые ветром частицы.[33] По мере осаждения и захоронения органических веществ более легко усваиваемые соединения истощаются в результате микробного окисления, оставляя более стойкие соединения. Таким образом, энергия, доступная для жизни, уменьшается. В нескольких верхних метрах скорость метаболизма снижается на 2–3 порядка, а количество клеток в осадочной колонке уменьшается с глубиной.[48]
Осадки образуют слои с разными условиями для жизни. В верхних 5–10 сантиметрах животные зарываются, перерабатывая осадок и расширяя границу раздела осадок-вода. Вода несет в себе кислород, свежие органические вещества и растворенные метаболиты, что приводит к неоднородной среде с обильными питательными веществами. Ниже залегающего слоя находится слой, в котором преобладает сульфатредукция. Ниже этого анаэробного восстановления метана способствует сульфат в сульфатно-метановая переходная зона (СМТЗ). Как только сульфаты истощаются, образование метана берет на себя.[48] Глубина химических зон зависит от скорости осаждения органических веществ. Там, где это происходит быстро, кислород быстро поглощается по мере потребления органических веществ; там, где медленно, кислород может оставаться намного глубже из-за недостатка питательных веществ для окисления.[48]
Среды обитания океанических отложений можно разделить на субдукция зоны, абиссальные равнины, и пассивная наценка. В зоне субдукции, где одна плита ныряет под другую, имеет тенденцию формироваться толстый клин осадка. Сначала в осадке от 50 до 60 процентов. пористость; при сжатии жидкости вытесняются с образованием холодные просачивания или же газовые гидраты.
Абиссальные равнины - это регион между континентальные окраины и срединно-океанические хребты, обычно на глубине менее 4 км. Поверхность океана очень бедна питательными веществами, такими как нитраты, фосфаты и железо, что ограничивает рост фитопланктон; это приводит к низкой скорости осаждения.[52] Осадок, как правило, очень беден питательными веществами, поэтому не весь кислород потребляется; кислород был обнаружен вплоть до нижележащая порода. В таких средах клетки в основном либо строго аэробный или же факультативный анаэробный (используя кислород, если он есть, но может переключаться на другие акцепторы электронов в его отсутствие[65]) и они являются гетеротрофный (не первичные производители). К ним относятся протеобактерии, хлорофлекси, археи морской группы II и литоавтотрофы в филуме Thaumarchaeota. Грибы разнообразны, в том числе представители Аскомикота и Базидиомицеты филы, а также дрожжи[52]
Пассивные окраины (континентальные шельфы и склоны) находятся под относительно мелководьем. Апвеллинг выносит на поверхность воду, богатую питательными веществами, стимулируя обильный рост фитопланктона, который затем оседает на дно (явление, известное как биологический насос ).[52] Таким образом, в отложениях много органического материала, и весь кислород расходуется на его потребление. У них очень стабильные профили температуры и давления.[36] Население микробов на порядки больше, чем на абиссальных равнинах. Это включает в себя строгие анаэробы включая членов группы Chloroflexi, "Ca. Атрибактерии », сульфатредуцирующие бактерии, и ферментеры, метаногены и метанотрофы в архее. Грибы менее разнообразны, чем на абиссальных равнинах, в основном включают Ascomycota и дрожжи. Вирусы в Inoviridae, Siphoviridae и Lipothrixviridae семьи были идентифицированы.[52]
Горные породы

Кора океана образуется в срединно-океанические хребты и удаляется субдукцией. Верхние полкилометра или около того представляют собой серию базальтовых потоков, и только этот слой имеет достаточную пористость и проницаемость, чтобы позволить течь жидкости. Менее пригодны для жизни слои обшитые дамбой и габбро под.[36]
Срединно-океанические хребты - это горячая, быстро меняющаяся среда с крутым вертикальным температурным градиентом, поэтому жизнь может существовать только в верхних нескольких метрах. Высокотемпературные взаимодействия между водой и горной породой уменьшают содержание сульфатов, производя большое количество сульфидов, которые служат источниками энергии; они также очищают породу от металлов, которые могут быть источниками энергии или токсичными. Помимо дегазации из магмы, при взаимодействии с водой также образуется много метана и водорода. Бурение еще не проводилось, поэтому информация о микробах поступает из образцов гидротермальных жидкостей, выходящих из жерл.[36]
Примерно в 5 километрах от оси хребта, когда земной коре около 1 миллиона лет, начинаются фланги хребта. Был характеризован гидротермальная циркуляция, их возраст составляет около 80 миллионов лет. Эта циркуляция вызвана скрытым теплом от охлаждения коры, которое нагревает морскую воду и прогоняет ее вверх через более проницаемые породы. Источниками энергии являются изменения горных пород, некоторые из которых опосредованы живыми организмами. В молодой коре много железа и серы. Покрытие осадка замедляет охлаждение и уменьшает поток воды. Существует мало свидетельств активности микробов в более старой (более 10 миллионов лет) корке.[36]
Вблизи зон субдукции вулканы могут образовываться в островные дуги и задняя дуга регионы. Подводящая плита выделяет летучие вещества и растворенные вещества в эти вулканы, в результате чего образуются кислые флюиды с более высокими концентрациями газов и металлов, чем в срединно-океаническом хребте. Он также выделяет воду, которая может смешиваться с материалом мантии, образуя серпентинит. Когда горячая точка вулканы расположены посреди океанических плит, они создают проницаемые и пористые базальты с более высокими концентрациями газа, чем на срединно-океанических хребтах. Гидротермальные флюиды более холодные и имеют более низкое содержание сульфидов. Бактерии, окисляющие железо, создают обширные отложения оксидов железа.[36]
Грунтовая вода
Микроорганизмы обитают в трещинах, отверстиях и пустотах внутри отложений и горных пород. Такое пустое пространство обеспечивает микроорганизмы водой и растворенными питательными веществами. Обратите внимание, что по мере увеличения глубины в глубине становится меньше питательных веществ. грунтовая вода поскольку питательные вещества постоянно потребляются микроорганизмами. По мере увеличения глубины осадка становится больше. компактный и между ними меньше места минеральная зерна. В результате на единицу объема приходится меньше поровой воды. Окружающая среда становится все суше и суше, когда отложения переходят в породы. На этом этапе вода также может быть ограничивающим фактором для глубинной биосферы.[30]
Континенты
Континенты имеют сложную историю и большое разнообразие горных пород, отложений и почв; климат на поверхности, профили температуры и гидрология также различаются. Большая часть информации о подземной жизни поступает из небольшого числа участков отбора проб, которые в основном находятся в Северной Америке. За исключением ледяных кернов, плотность ячеек резко снижается с глубиной, уменьшаясь на несколько порядков. В верхних 1-2 метрах почвы организмы зависят от кислорода и гетеротрофы, в зависимости от расщепления органического углерода для их питания, и их снижение плотности аналогично падению плотности органического материала. Ниже этого нет корреляции, хотя как плотность клеток, так и органическое содержание снижаются еще на пять. порядки величины или около того (напротив, в океанических отложениях есть корреляция). Увеличение глубины, температуры и солености действительно коррелирует с уменьшением числа клеток, хотя скорость сильно зависит от типа коры и скорости подпитка подземных вод.[54]
Микробы были обнаружены в осадочных породах на глубине до 3 километров, самая глубокая из них. Есть много разнообразия, хотя самые глубокие, как правило, железо (III) - или сульфатредуцирующие бактерии, которые используют ферментацию и могут процветать при высокой температуре и солености. Еще более солеустойчивый галофилы были найдены в глубоких соляных отложениях, и такие отложения встречаются по всему миру.[66] В 2019 году были обнаружены микробные организмы, живущие на глубине 2400 метров под поверхностью, дышащие серой и поедающие камни, такие как пирит, в качестве обычного источника пищи.[67][68][69] Открытие произошло в старейшей известной воде на Земле.[70]
Люди получали доступ к глубоким водоносным горизонтам в вулканических породах для различных целей, включая добычу подземных вод, добычу полезных ископаемых и хранение опасных отходов. В большинстве или во всех этих водоносных горизонтах обитают микробы. На всех проверенных объектах обнаружены водород, метан и углекислый газ.[66] Сообщества прокариот, основанные на водороде, также были обнаружены в горячих источниках и гидротермальных системах. Было предложено множество механизмов производства водорода, некоторые из которых не зависят от фотосинтеза.[71]
Экология

Один вид бактерий "Candidatus Desulforudis audaxviator ", является первым из известных, составляющих целую экосистему отдельно.[9] Он был найден на 2,8 км под землей в золотом руднике недалеко от Йоханнесбург, Южная Африка. В щелочной вода с температурой около 60 ° C, не имеющая доступа к кислороду, получает энергию за счет восстановления сульфата, его азота из молекул аммиака и ионов аммония, а углерода из диоксида углерода или формиата.[72][73]
В других экосистемах есть несколько взаимозависимых видов. Их можно разделить на автотрофы, которые получают энергию из неживых источников, и гетеротрофы, которые питаются автотрофами или их останками. Некоторые организмы участвуют в синтрофия, где один организм живет за счет побочных продуктов метаболической активности другого. На поверхности большинство автотрофов используют фотосинтез, но там, где нет света, хемоавтотрофы использовать химическую энергию.[74]
В морских отложениях, где доступен кислород, основная группа хемоавтотрофов - это таумархеи, окисляющие аммиак. Он поддерживает 19% гетеротрофного производства. В некоторых средах, таких как абиссальные отложения Тихого океана, запас аммиака уменьшается с глубиной; но в других средах содержание аммиака увеличивается, потому что гетеротрофные бактерии, живущие на органическом материале, реминерализуют аммиак. Эта взаимозависимость гетеротрофных бактерий и Thaumarchaeota является примером синтрофии. Однако некоторые таумархеи миксотрофный, способные использовать как органические вещества, так и диоксид углерода в качестве углерода.[52]
В бескислородных отложениях водород является важным «съедобным». Члены Chloroflexi черпают из него энергию для производства ацетат за счет уменьшения углекислого газа или органических веществ (процесс, известный как ацетогенез ). Продукция Bacteroidetes, восстанавливающая металл и сахарное брожение пропионат, среди других соединений, и это ферментируется "Ca. Атрибактерии "производят водород. В верхних отложениях сульфатредуцирующие бактерии поглощают большую часть водорода, в то время как в нижних отложениях сульфат истощается, и преобладают метаногены. В зоне перехода сульфат-метан (SMTZ) анаэробные метанотрофные (ANME) археи образуют консорциумы с сульфатредуцирующими бактериями.[52][48]
Смотрите также
Примечания
Рекомендации
- ^ а б c d е ж Hoehler, Tori M .; Йоргенсен, Бо Баркер (16 января 2013 г.). «Микробная жизнь при крайнем ограничении энергии». Обзоры природы Микробиология. 11 (2): 83–94. Дои:10.1038 / nrmicro2939. HDL:2060/20150018056. PMID 23321532.
- ^ Шипперс, А. (2015). «Глубокая биосфера». В Harff, J .; Мешеде, М .; Petersen, S .; Thiede, J. (ред.). Энциклопедия морских геонаук. Дордрехт: Спрингер. ISBN 978-94-007-6644-0.
- ^ а б c Бар-Он, Йинон М .; Филлипс, Роб; Майло, Рон (19 июня 2018 г.). «Распределение биомассы на Земле». Труды Национальной академии наук. 115 (25): 6506–6511. Дои:10.1073 / pnas.1711842115. ЧВК 6016768. PMID 29784790.
- ^ а б c Эдвардс, Катрина (2 сентября 2011 г.). «Северный пруд: в поисках внеземной жизни». Сеть блогов Scientific American. Получено 18 января 2019.
- ^ Джадсон, Оливия (10 июня 2008 г.). "Знакомство с инопланетянами". Автор мнения. Получено 18 января 2019.
- ^ Элли, Уильям М. (1993). Региональное качество грунтовых вод. Джон Вили и сыновья. п. 182. ISBN 9780471284536.
- ^ Уорд, Питер Д .; Браунли, Дональд (2006). Редкая земля: почему сложная жизнь необычна во Вселенной (Пбк. Ред.). Коперник. С. 7–12. ISBN 9780387218489.
- ^ а б c Онстотт 2016, Глава 1
- ^ а б c d Ли Маскарелли, Аманда (11 июня 2009 г.). "Низкий уровень жизни". Природа. 459 (7248): 770–773. Дои:10.1038 / 459770a.
- ^ а б c d Эдвардс, Катрина Дж .; Беккер, Кейр; Колвелл, Фредерик (30 мая 2012 г.). «Глубокая биосфера темной энергии: внутриземная жизнь на Земле». Ежегодный обзор наук о Земле и планетах. 40 (1): 551–568. Дои:10.1146 / аннурьев-земля-042711-105500.
- ^ а б Фредриксон, Джеймс К .; Онстотт, Таллис С. (1996). «Микробы глубоко внутри Земли». Scientific American. 275 (4): 68–73. Дои:10.1038 / scientificamerican1096-68. JSTOR 24993405. PMID 8797299.
- ^ а б c d е ж грамм час я Colwell, F. S .; Д'Хонд, С. (13 февраля 2013 г.). «Природа и масштабы глубинной биосферы». Обзоры по минералогии и геохимии. 75 (1): 547–574. Дои:10.2138 / RMG.2013.75.17.
- ^ а б Онстотт 2016, Глава 6
- ^ «Этап 201: Контроль микробных сообществ в глубоко погребенных отложениях, восточная часть Экваториальной части Тихого океана и участки на окраине Перу 1225–1231». Программа морского бурения. Получено 20 июля 2019.
- ^ а б c d е Deep Carbon Observatory (2019). Обсерватория глубинного углерода: десятилетие открытий. Вашингтон, округ Колумбия. Дои:10.17863 / CAM.44064. Получено 13 декабря 2019.
- ^ Кифт, Т .; Phelps, T .; Фредриксон, Дж. (2007). «66. Бурение, бурение кернов и отбор проб подземных сред». В Hurst, C .; Crawford, R .; Garland, J .; Lipson, D .; Миллс, А .; Стеценбах, Л. (ред.). Руководство по экологической микробиологии (3-е изд.). Вашингтон, округ Колумбия: ASM Press. Дои:10.1128 / 9781555815882.ch66. ISBN 9781555815882.
- ^ Gihring, T. M .; Moser, D. P .; Lin, L.-H .; Дэвидсон, М .; Onstott, T. C .; Morgan, L .; Milleson, M .; Kieft, T. L .; Тримарко, Э. (1 сентября 2006 г.). «Распределение микробных таксонов в подземных водах щита Калахари, Южная Африка». Журнал геомикробиологии. 23 (6): 415–430. Дои:10.1080/01490450600875696. ISSN 0149-0451.
- ^ Итаваара, Мерджа; Ахонен, Лассе; Нумминен, Микко; Зольберг, Элина; Киетявяйнен, Риикка; Миеттинен, Ханна (2015). «Состав микробиома и геохимические характеристики глубинной подземной среды высокого давления, рудник Пюхясалми, Финляндия». Границы микробиологии. 6. Дои:10.3389 / fmicb.2015.01203. ISSN 1664-302X.
- ^ Мангельсдорф, Кай; Каллмейер, Йенс (10 сентября 2010 г.). «Интеграция глубинных исследований биосферы в международную континентальную программу научного бурения». Научное бурение (10 сентября 2010 г.). Дои:10.2204 / iodp.sd.10.0.2010.
- ^ "ПРОБКИ". Ocean Networks Canada. 15 октября 2012 г.. Получено 18 июля 2019.
- ^ Беккер, Кейр; Дэвис, Эрл Э. (31 октября 2005 г.). «Обзор проектов и операций CORK во время программы морского бурения». Труды Комплексной программы морского бурения. Труды IODP. 301. Дои:10.2204 / iodp.proc.301.104.2005.
- ^ «Список инструментов для пакеров и расходомеров». Обзор технических средств и оборудования программы морского бурения. Программа морского бурения. Получено 23 августа 2019.
- ^ Кулин, Ян; Ридель, Майкл; Клаус, Адам (2013). «Простой инструмент с кабелем для измерения параметров на месте (SCIMPI) и замена CORK в отверстии 858G». Научный проспект ИОДП. 341S. Дои:10.2204 / iodp.sp.341S.2013.
- ^ Пуркамо, Лотта; Бомберг, Малин; Нюиссонен, Мари; Кукконен, Ильмо; Ахонен, Лассе; Киетявяйнен, Риикка; Итаваара, Мерджа (2013). «Расчленение глубинной биосферы: извлечение аутентичных микробных сообществ из изолированных пакером глубоких кристаллических зон трещиноватости коренных пород». FEMS Microbiology Ecology. 85 (2): 324–337. Дои:10.1111/1574-6941.12126. ISSN 1574-6941. PMID 23560597.
- ^ Нурми, Пекка А .; Кукконен, Ильмо Т. (1986). «Новый метод отбора проб воды и газа из глубоких скважин». Канадский журнал наук о Земле. 23: 1540–1454.
- ^ Киетявяйнен, Риикка; Ахонен, Лассе; Кукконен, Ильмо Т .; Хендрикссон, Нина; Нюиссонен, Мари; Итаваара, Мерджа (1 мая 2013 г.). «Характеристика и изотопная эволюция соленых вод глубокой буровой скважины Оутокумпу, Финляндия - Последствия для происхождения воды и глубинной земной биосферы». Прикладная геохимия. Специальный выпуск, посвященный 9-му Международному симпозиуму по прикладной геохимии изотопов (AIG9), Таррагона, Испания, сентябрь 2011 г. 32: 37–51. Дои:10.1016 / j.apgeochem.2012.10.013. ISSN 0883-2927.
- ^ Haggerty, R .; Schroth, M.H .; Исток, J.D. (март 1998 г.). «Упрощенный метод анализа данных« push-Pull »для определения коэффициентов скорости реакции in situ». Грунтовые воды. 36 (2): 314–324. Дои:10.1111 / j.1745-6584.1998.tb01097.x.
- ^ Сандерс, Роберт (5 мая 2005 г.). «Протеомика приближает исследователей к пониманию микробов, которые производят кислотный дренаж шахт» (Пресс-релиз). Калифорнийский университет в Беркли. Получено 19 июля 2019.
- ^ а б Орелл, Альваро; Фрелс, Сабрина; Альберс, Соня-Верена (8 сентября 2013 г.). «Архейские биопленки: великое неизведанное». Ежегодный обзор микробиологии. 67 (1): 337–354. Дои:10.1146 / annurev-micro-092412-155616.
- ^ а б c d е ж грамм «Микробная жизнь глубоко под морским дном - история о том, как не сдаваться». Границы для молодых умов. Дои:10.3389 / frym.2020.00070. Получено 9 июн 2020.
- ^ "Экспедиция в затерянный город 2005". Ocean Explorer. Национальное управление океанических и атмосферных исследований. Получено 11 ноября 2019.
- ^ Поль, Уолтер (2011). Экономическая геология: принципы и практика. Вайли. п. 89. ISBN 9781444394863.
- ^ а б c d е ж грамм Orcutt, B.N .; Sylvan, J. B .; Knab, N.J .; Эдвардс, К. Дж. (6 июня 2011 г.). «Микробная экология Темного океана над, на и под морским дном». Обзоры микробиологии и молекулярной биологии. 75 (2): 361–422. Дои:10.1128 / MMBR.00039-10. PMID 21646433.
- ^ Нилсон, Кеннет Х. (март 2003 г.). «Использование аппетитов микробов для восстановления». Природа Биотехнологии. 21 (3): 243–244. Дои:10.1038 / nbt0303-243. PMID 12610569.
- ^ Йоргенсен, Бо Баркер; Боэтиус, Антье (октябрь 2007 г.). «Пир и голод - микробная жизнь на глубоководном дне». Обзоры природы Микробиология. 5 (10): 770–781. Дои:10.1038 / nrmicro1745.
- ^ а б c d е ж грамм час я j k л Шренк, Мэтью О .; Huber, Julie A .; Эдвардс, Катрина Дж. (Январь 2010 г.). «Микробные провинции на дне моря». Ежегодный обзор морской науки. 2 (1): 279–304. Дои:10.1146 / аннурьев-марин-120308-081000. PMID 21141666.
- ^ а б c d Ротшильд, Линн Дж .; Манчинелли, Рокко Л. (февраль 2001 г.). «Жизнь в экстремальных условиях». В Ямагиши, Акихико; Какегава, Такеши; Усуи, Томхиро (ред.). Астробиология: от истоков жизни до поисков внеземного разума. Природа. 409. Сингапур: Спрингер. С. 1092–1101. Дои:10.1038/35059215. ISBN 978-981-13-3639-3.
- ^ Карио, Анаис; Оливер, Джина С.; Роджерс, Карин Л. (4 сентября 2019 г.). «Изучение глубоководной морской биосферы: проблемы, инновации и возможности». Границы науки о Земле. 7. Дои:10.3389 / feart.2019.00225.
- ^ «Глубоководные микробы предпочитают образ жизни с высоким давлением». Глубокая углеродная обсерватория. 23 июля 2019 г.. Получено 15 ноября 2019.
- ^ Тамбурини, Кристиан; Бутриф, Мехди; Гарель, Марк; Colwell, Rita R .; Деминг, Джоди В. (май 2013 г.). «Прокариотические реакции на гидростатическое давление в океане - обзор» (PDF). Экологическая микробиология. 15 (5): 1262–1274. Дои:10.1111/1462-2920.12084.
- ^ "PUSH для микробиологии высокого давления". Глубокая углеродная обсерватория. 30 сентября 2019 г.. Получено 15 ноября 2019.
- ^ а б Хойер, Верена; Рычаг, Марк; Мороно, Юки; Теске, Андреас (1 марта 2019 г.). «Пределы жизни и биосферы в недрах Земли». Океанография. 32 (1): 208–211. Дои:10.5670 / oceanog.2019.147.
- ^ Маккей, К. П. (9 июня 2014 г.). «Требования и ограничения для жизни в контексте экзопланет». Труды Национальной академии наук. 111 (35): 12628–12633. Дои:10.1073 / pnas.1304212111.
- ^ Такай, Кен (2019). «Пределы земной жизни и биосферы». Астробиология. С. 323–344. Дои:10.1007/978-981-13-3639-3_20. ISBN 978-981-13-3638-6.
- ^ а б c d е ж Colman, Daniel R .; Пудель, Сародж; Марки, Блейк У .; Бойд, Эрик С .; Спир, Джон Р. (3 июля 2017 г.). «Глубокая горячая биосфера: двадцать пять лет ретроспективы». Труды Национальной академии наук. 114 (27): 6895–6903. Дои:10.1073 / pnas.1701266114.
- ^ Биддл, Дженнифер Ф .; Сильван, Джейсон Б.; Brazelton, Уильям Дж .; Талли, Бенджамин Дж .; Эдвардс, Катрина Дж .; Мойер, Крейг Л .; Хедельберг, Джон Ф .; Нельсон, Уильям К. (2012). «Перспективы изучения эволюции в глубинной биосфере». Границы микробиологии. 2: 285. Дои:10.3389 / fmicb.2011.00285.
- ^ а б c Коллинз, Терри; Пратт, Кэти (10 декабря 2018 г.). «Жизнь в глубинах Земли содержит от 15 до 23 миллиардов тонн углерода - в сотни раз больше, чем у людей» (Пресс-релиз). Глубокая углеродная обсерватория. Получено 14 июля 2019.
- ^ а б c d е ж грамм Петро, К; Старнавский, П; Шрамм, А; Kjeldsen, KU (12 июня 2017 г.). «Сборка микробного сообщества в морских отложениях». Экология водных микробов. 79 (3): 177–195. Дои:10.3354 / ame01826.
- ^ а б Паркс, Р. Джон; Крэгг, Барри; Руссель, Эрван; Вебстер, Гордон; Весман, Эндрю; Сасс, Хенрик (июнь 2014 г.). «Обзор прокариотических популяций и процессов в донных отложениях, включая биосферу: геосферные взаимодействия». Морская геология. 352: 409–425. Дои:10.1016 / j.margeo.2014.02.009.
- ^ Hug, Laura A .; Бейкер, Бретт Дж .; Анантараман, Картик; Браун, Кристофер Т .; Probst, Alexander J .; Кастель, Синди Дж .; Баттерфилд, Кристина Н .; Hernsdorf, Alex W .; Амано, Юки; Исэ, Котаро; Сузуки, Йохей; Дудек, Наташа; Relman, David A .; Finstad, Kari M .; Амундсон, Рональд; Томас, Брайан Ч .; Банфилд, Джиллиан Ф. (11 апреля 2016 г.). «Новый взгляд на древо жизни». Природная микробиология. 1 (5). Дои:10.1038 / NMICROBIOL.2016.48.
- ^ Ли, Юнг Ми; Хван, Кюин; Ли, Чжэ Иль; Ким, Минчол; Хван, Чон Ён; Но, Хён-Джу; Чой, Хаккьюм; Ли, Хонг Кум; Чун, Чонгсик; Хонг, Сун Гю; Шин, Сын Чхоль (29 ноября 2018 г.). "Геномное понимание преобладания линии происхождения Candidate Phylum Atribacteria JS1 в морских отложениях". Границы микробиологии. 9. Дои:10.3389 / fmicb.2018.02909.
- ^ а б c d е ж грамм Орси, Уильям Д. (2 июля 2018 г.). «Экология и эволюция морских и подводных микробных сообществ». Обзоры природы Микробиология. 16 (11): 671–683. Дои:10.1038 / s41579-018-0046-8. PMID 29967420.
- ^ Маклауд, Фрейзер; С. Киндлер, Гарет; Лун Вонг, достопочтенный; Чен, Рэй; П. Бернс, Брендан (2019). «Археи Асгарда: разнообразие, функции и эволюционные последствия для ряда микробиомов». AIMS Microbiology. 5 (1): 48–61. Дои:10.3934 / microbiol.2019.1.48.
- ^ а б Magnabosco, C .; Lin, L.-H .; Dong, H .; Бомберг, М .; Ghiorse, W .; Stan-Lotter, H .; Pedersen, K .; Kieft, T. L .; van Heerden, E .; Онстотт, Т.К. (24 сентября 2018 г.). «Биомасса и биоразнообразие континентальных недр». Природа Геонауки. 11 (10): 707–717. Дои:10.1038 / s41561-018-0221-6.
- ^ Мошер, Дэйв (2 июня 2011 г.). "Новый" дьявольский червь "- самое глубокоживущее животное". National Geographic News. Получено 2 июля 2019.
- ^ «Глубинная жизнь не ограничивается микробами: землетрясения перемещают наземных животных в глубины» (Пресс-релиз). Глубокая углеродная обсерватория. 4 марта 2019 г.. Получено 14 июля 2019.
- ^ Итаваара, Мерджа; Викман, Минна; Салавирта, Хейкки; Нюиссонен, Мари; Миеттинен, Ханна; Бомберг, Малин; Зольберг, Элина (2015). «Выявление неисследованных грибных сообществ в глубоких грунтовых водах в зонах разломов кристаллической коренной породы в Олкилуото, Финляндия». Границы микробиологии. 6. Дои:10.3389 / fmicb.2015.00573. ISSN 1664-302X.
- ^ Бомберг, Малин; Итаваара, Мерджа; Кукконен, Ильмо; Зольберг, Элина; Миеттинен, Ханна; Киетявяйнен, Риикка; Пуркамо, Лотта (1 августа 2018 г.). «Разнообразие и функциональность сообществ архей, бактерий и грибов в глубоких архейских коренных подземных водах». FEMS Microbiology Ecology. 94 (8). Дои:10.1093 / фемсек / fiy116. ISSN 0168-6496.
- ^ «Грибы - ключевые игроки глубинной биосферы». ScienceDaily. Получено 22 августа 2019.
- ^ Borgonie, G .; Linage-Alvarez, B .; Ojo, A.O .; Mundle, S.O.C .; Freese, L B .; Van Rooyen, C .; Кулойо, О .; Albertyn, J .; Pohl, C .; Cason, E.D .; Vermeulen, J .; Pienaar, C .; Litthauer, D .; Van Niekerk, H .; Van Eeden, J .; Лоллар, Б. Шервуд .; Onstott, T. C .; Ван Херден, Э. (24 ноября 2015 г.). «Эукариотические оппортунисты доминируют в глубоководной биосфере Южной Африки». Nature Communications. 6 (1): 8952. Дои:10.1038 / ncomms9952. ЧВК 4673884. PMID 26597082.
- ^ Равиндран, Сандип (29 февраля 2016 г.). «Внутренняя Земля изобилует экзотическими формами жизни». Смитсоновский институт. Получено 14 июля 2019.
- ^ Иварссон, М .; Bengtson, S .; Нойбек, А. (апрель 2016 г.). «Огненная океаническая кора - самая большая среда обитания грибов на Земле?». Грибковая экология. 20: 249–255. Дои:10.1016 / j.funeco.2016.01.009.
- ^ Голд, Т. (1 июля 1992 г.). «Глубокая горячая биосфера». Труды Национальной академии наук. 89 (13): 6045–6049. Дои:10.1073 / пнас.89.13.6045. ISSN 0027-8424. ЧВК 49434. PMID 1631089.
- ^ Амос, Джонатан (10 декабря 2018 г.). «Огромный масштаб жизни под нашими ногами». Новости BBC. Получено 14 июля 2019.
- ^ Хентжес, Д. Дж. (1996). «17. Анаэробы: общая характеристика». В Бароне, С. (ред.). Медицинская микробиология (4-е изд.). Медицинский филиал Техасского университета в Галвестоне. Получено 19 ноября 2019.
- ^ а б Педерсен, К. (апрель 2000 г.). «Исследование глубинных внутриземных микробов: современные перспективы». Письма о микробиологии FEMS. 185 (1): 9–16. Дои:10.1111 / j.1574-6968.2000.tb09033.x.
- ^ Lollar, Garnet S .; Уорр, Оливер; Рассказывая, Джон; Osburn, Magdalena R .; Лоллар, Барбара Шервуд (18 июля 2019 г.). "'«Следуй за водой»: гидрогеохимические ограничения на микробные исследования. 2,4 км под поверхностью в обсерватории глубинных флюидов и глубинной жизни Кидд-Крик ». Журнал геомикробиологии. 36 (10): 859–872. Дои:10.1080/01490451.2019.1641770.
- ^ «Самые старые подземные воды в мире поддерживают жизнь благодаря химическому составу воды и горных пород». Глубокая углеродная обсерватория. 29 июля 2019 г.. Получено 15 ноября 2019.
- ^ Пауэлл, Кори С. (7 сентября 2019 г.). "Странные формы жизни, найденные глубоко в шахте, указывают на огромные" подземные Галапагосы ".'". Новости NBC. Получено 15 ноября 2019.
- ^ Ромулд, Мэгги (14 декабря 2016 г.). «Древнейшая вода на Земле, найденная глубоко под канадским щитом». Исследователь науки. Получено 15 ноября 2019.
- ^ Паркс, Р. Джон; Крэгг, Барри; Руссель, Эрван; Вебстер, Гордон; Весман, Эндрю; Сасс, Хенрик (июнь 2014 г.). «Обзор прокариотических популяций и процессов в донных отложениях, включая биосферу: геосферные взаимодействия». Морская геология. 352: 409–425. Дои:10.1016 / j.margeo.2014.02.009.
- ^ Старр, Лаура (9 октября 2008 г.). «Один - самый одинокий номер для бактерии, обитающей в шахте». Природа. Дои:10.1038 / новости.2008.1160.
- ^ Министерство энергетики / Национальная лаборатория Лоуренса Беркли (10 октября 2008 г.). «Путешествие к центру Земли: единственный в своем роде микроорганизм живет один». ScienceDaily. Получено 21 ноября 2019.
- ^ Chapman, J. L .; Рейсс, Майкл Дж. (1999). Экология: принципы и приложения (2-е изд.). Издательство Кембриджского университета. С. 8, 120–121. ISBN 9780521588027.
дальнейшее чтение
- Абэ, Шиге (10 октября 2008 г.). «Жизнь без Солнца». Астробиология в НАСА. Получено 16 июля 2019.
- «Аминокислотный метаболизм способствует развитию сообществ гидроразрыва». Глубокая углеродная обсерватория (Пресс-релиз). 13 августа 2018 г.. Получено 3 сентября 2019.
- Биддл, Дженнифер Ф. (2012). «Перспективы изучения эволюции в глубинной биосфере». Границы микробиологии. 2: 285. Дои:10.3389 / fmicb.2011.00285.
- Бомберг, Малин; Ахонен, Лассе (2017). Геомикробы: жизнь в глубоких земных недрах. Frontiers Media SA. ISBN 9782889451791.
- Брэдли, Джеймс А .; Amend, Ян П .; ЛаРоу, Дуглас Э. (январь 2019 г.). «Выживание наименьшего количества: покой микробов и поддержание в морских отложениях в глубоких временах». Геобиология. 17 (1): 43–59. Дои:10.1111 / gbi.12313. ЧВК 6585783. PMID 30248245.
- Брахич, Екатерина (9 декабря 2013 г.). «19 супербактерий, управляющих скрытыми глубинами Земли». Новый ученый. Получено 3 сентября 2019.
- Браун (директор), Майкл; Сантелл (Сюжет), Ад Люсьен (3 декабря 2013 г.). Северный пруд: В поисках инопланетян (Видео). Центр исследований темной энергии биосферы. Получено 10 июля 2019. (IMDb )
- Д'Хонд, С. (15 марта 2002 г.). «Метаболическая активность подземной жизни в глубоководных отложениях». Наука. 295 (5562): 2067–2070. Дои:10.1126 / science.1064878.
- Фрейзер, Дженнифер (23 января 2019 г.). «Внутри Земли микробы приближаются к бессмертию». Сеть блогов Scientific American. Получено 5 июля 2019.
- Гхош, Тиа (29 декабря 2013 г.). «Что скрывается под землей: крошечные организмы процветают под поверхностью Земли». Живая наука. Получено 16 июля 2019.
- Гулд, Стивен Джей (13 ноября 1996 г.). «Планета бактерий». Washington Post Horizon. 119: 344. Получено 10 июля 2019.
- Хигнетт, Кэтрин (18 декабря 2017 г.). "Ученые искали жизнь на Марсе не в том месте?". Newsweek. Получено 18 января 2019.
- Hinrichs, K.-U .; Инагаки, Ф. (11 октября 2012 г.). «Сокращение глубин биосферы». Наука. 338 (6104): 204–205. Дои:10.1126 / science.1229296.
- Каллмейер, Йенс; Вагнер, Дирк (2014). Микробная жизнь глубокой биосферы. Вальтер Де Грюйтер. ISBN 9783110370676.
- Лейтч, Кармен (10 декабря 2018 г.). «Глубокая биосфера Земли изобилует жизнью». LabRoots. Получено 18 января 2019.
- Рычаг, Марка А .; Роджерс, Карин Л .; Ллойд, Карен Г .; Оверманн, Йорг; Шинк, Бернхард; Thauer, Rudolf K .; Hoehler, Tori M .; Йоргенсен, Бо Баркер; Джудичи-Ортикони, Мария-Тереза (сентябрь 2015 г.). «Жизнь в условиях экстремального ограничения энергии: синтез лабораторных и полевых исследований». Обзор микробиологии FEMS. 39 (5): 688–728. Дои:10.1093 / femsre / fuv020. PMID 25994609.
- Нилсон, Кеннет Х. (март 2003 г.). «Использование аппетитов микробов для восстановления». Природа Биотехнологии. 21 (3): 243–244. Дои:10.1038 / nbt0303-243. PMID 12610569.
- Онстотт, Таллис С. (2014). «Глубинная подповерхностная микробиология». Энциклопедия астробиологии. Берлин, Гейдельберг: Springer Berlin Heidelberg. С. 1–4. Дои:10.1007/978-3-642-27833-4_573-3. ISBN 9783642278334.
- Онстотт, Таллис К. (2016). Глубокая жизнь. Издательство Принстонского университета. ISBN 9781400884247.CS1 maint: ref = harv (связь)
- Теске, Андреас; Соренсен, Кетил Б. (8 ноября 2007 г.). «Невыводимые археи в глубоководных морских отложениях: мы их всех поймали?». Журнал ISME. 2 (1): 3–18. Дои:10.1038 / ismej.2007.90. PMID 18180743.
- Трембат-Райхерт, Элизабет; Мороно, Юки; Иджири, Акира; Хосино, Тацухико; Доусон, Кэтрин С .; Инагаки, Фумио; Сирота, Виктория Дж. (31 октября 2017 г.). «Использование метиловых соединений и медленный рост характеризуют микробную жизнь в угольных и сланцевых пластах глубиной 2 км». Труды Национальной академии наук. 114 (44): E9206 – E9215. Дои:10.1073 / pnas.1707525114. PMID 29078310.
- Уоттс, Джонатан (10 декабря 2018 г.). «Ученые идентифицируют обширную подземную экосистему, содержащую миллиарды микроорганизмов». Хранитель. Получено 18 января 2019.