Термогенин - Thermogenin




Термогенин (называется разобщающий белок его первооткрыватели и теперь известный как разобщающий белок 1, или UCP1)[5] это митохондриальный носитель белок содержится в коричневая жировая ткань (ЛЕТУЧАЯ МЫШЬ). Он используется для выработки тепла за счет отсутствия дрожи термогенез, и вносит количественно важный вклад в противодействие потере тепла у младенцев, которая в противном случае произошла бы из-за их высокого отношения площади поверхности к объему.
Механизм
UCP представляют собой трансмембранные белки, которые уменьшают протонный градиент, образующийся при окислительном фосфорилировании. Они делают это за счет увеличения проницаемости внутренней митохондриальной мембраны, позволяя протонам, которые были накачаны в межмембранное пространство, возвращаться в митохондриальный матрикс. Выделение тепла в буром жире, опосредованное UCP1, разъединяет дыхательную цепь, обеспечивая быстрое окисление субстрата с низкой скоростью производства АТФ. UCP1 связан с другими переносчиками митохондриальных метаболитов, такими как транслокатор адениновых нуклеотидов, протонный канал в митохондриальная внутренняя мембрана что позволяет переносить протоны из митохондриальной межмембранное пространство к митохондриальный матрикс. UCP1 ограничен коричневая жировая ткань, где он обеспечивает механизм огромной способности ткани генерировать тепло.
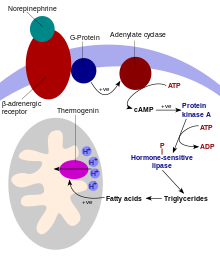
UCP1 активируется в коричневой жировой клетке жирными кислотами и ингибируется нуклеотидами.[6] Жирные кислоты высвобождаются с помощью следующего сигнального каскада: высвобождение окончаний симпатической нервной системы. Норэпинефрин на Бета-3-адренорецептор на плазматическая мембрана. Это активирует аденилилциклаза, который катализирует превращение АТФ в циклический AMP (лагерь). цАМФ активирует протеинкиназа А, вызывая освобождение его активных C-субъединиц от его регуляторных R-субъединиц. Активная протеинкиназа А, в свою очередь, фосфорилирует триацилглицерин липаза, тем самым активируя его. Липаза превращает триацилглицерины в свободные жирные кислоты, которые активируют UCP1, преодолевая ингибирование, вызываемое пуриновыми нуклеотидами (ВВП и ADP ). Во время завершения термогенеза термогенин инактивируется, а остаточные жирные кислоты удаляются путем окисления, позволяя клетке вернуться в нормальное состояние сохранения энергии.

UCP1 очень похож на белок-носитель АТФ / АДФ или транслокатор нуклеотидов аденина (МУРАВЕЙ ).[7] Предлагаемая модель переменного доступа для UCP1 основана на аналогичном механизме ANT.[8]. В субстрат поступает в полуоткрытый белок UCP1 из цитоплазматический сторону мембраны, белок закрывает цитоплазматическую сторону, так что субстрат заключен в белок, а затем матричная сторона белка открывается, позволяя субстрату высвобождаться в митохондриальный матрикс. Открытие и закрытие протеина достигается за счет затягивания и ослабления соляные мосты на мембранной поверхности белка. Обоснование этого моделирования UCP1 на ANT обнаруживается во многих консервативных остатках между двумя белками, которые активно участвуют в транспортировке субстрата через мембрану. Оба белка интегральные мембранные белки, локализованные на внутренней митохондриальной мембране, и они имеют похожую структуру солевых мостиков, пролин остатки и гидрофобный или же ароматный аминокислоты, которые могут закрываться или открываться в цитоплазматическом или матричном состоянии[7].
Эволюция
UCP1 экспрессируется в коричневой жировой ткани, которая функционально обнаруживается только в евтерийцы. Ген UCP1, или термогенин, вероятно, возник у предка современных позвоночные, но изначально не позволяли нашему предку позвоночных использовать отсутствие дрожи термогенез для тепла. Так было до тех пор, пока не было адаптивно выбранный для в плацентарный потомки млекопитающих этого общего предка, у которых UCP1 развил свою текущую функцию в коричневой жировой ткани, чтобы обеспечить дополнительное тепло[9]. В то время как UCP1 играет ключевую термогенную роль у плацентарных млекопитающих широкого спектра, особенно у млекопитающих с маленьким размером тела и у тех, кто впадает в спячку, ген UCP1 утратил функциональность у нескольких крупных клонов (например, лошади, слоны, морские коровы, киты и даманы ) и линии с низким уровнем метаболизма (например, панголины, броненосцы, ленивцы и муравьеды )[10]. Недавние открытия не тепловыделяющих ортологи UCP1 в рыбе и сумчатые, другие потомки предков современных позвоночных, показывают, что этот ген был передан всем современным позвоночным животным, но, кроме плацентарных млекопитающих, ни один из них не обладает способностью вырабатывать тепло.[11]. Это также предполагает, что UCP1 имел иную первоначальную цель, и на самом деле филогенетический анализ и анализ последовательности показывают, что UCP1, вероятно, является мутированной формой дикарбоксилатный носитель белок, адаптированный к термогенезу у плацентарных млекопитающих[12].
История
Исследователи 1960-х годов изучали коричневая жировая ткань, обнаружили, что в дополнение к выработке большего количества тепла, чем типично для других тканей, коричневая жировая ткань, по-видимому, замыкает или разъединяет дыхательную связь. [13]. Разобщающий белок 1 был открыт в 1978 году Дэвидом Николлсом, Вибеке Берсон и Джиллиан Хитон и показал, что он является белком, ответственным за этот эффект разобщения.[14]. Позднее UCP1 был впервые очищен в 1980 г. и впервые клонирован в 1988 г.[15][16]
Разъединение белка два (UCP2), гомолог UCP1, был идентифицирован в 1997 году. UCP2 локализуется в большом количестве тканей и, как полагают, участвует в регуляции активных форм кислорода (ROS). В последнее десятилетие были идентифицированы три дополнительных гомолога UCP1, включая UCP3, UCP4 и BMCP1 (также известный как UCP5).
Клиническая значимость
Способы доставки UCP1 в клетки с помощью терапии с переносом генов или методы его активации были важным направлением исследований в области лечения ожирения из-за их способности рассеивать избыточные метаболические запасы.[17]
Смотрите также
- 2,4-динитрофенол (Синтетический малая молекула протонный челнок с аналогичными эффектами)
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000109424 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000031710 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ген Entrez: UCP1, разобщающий белок 1 (митохондриальный, протонный переносчик)».
- ^ «Механизм зависимого от жирных кислот разобщения UCP1 в митохондриях бурого жира». Клетка. 151 (2): 400–413. 2012-10-12. Дои:10.1016 / j.cell.2012.09.010. ISSN 0092-8674.
- ^ а б Crichton, Paul G .; Ли, Ян; Кунджи, Эдмунд Р. С. (2017-03-01). «Молекулярные особенности разобщающего белка 1 поддерживают традиционный механизм митохондриального носителя». Биохимия. UCP1: 40 лет и старше. 134: 35–50. Дои:10.1016 / j.biochi.2016.12.016. ISSN 0300-9084. ЧВК 5395090. PMID 28057583.
- ^ Райан, Рене М .; Ванденберг, Роберт Дж. (2016-03-01). «Повышение модели переменного доступа». Структурная и молекулярная биология природы. 23 (3): 187–189. Дои:10.1038 / nsmb.3179. ISSN 1545-9985. PMID 26931415. S2CID 35913348.
- ^ Клингенспор, Мартин; Фромме, Тобиас; Хьюз, Дэвид А .; Манцке, Ларс; Полимеропулос, Элиас; Риман, Тобиас; Трженка, Магдалина; Хиршберг, Верена; Ястрох, Мартин (01.07.2008). «Древний взгляд на UCP1». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 15-я Европейская конференция по биоэнергетике 2008 г. 1777 (7): 637–641. Дои:10.1016 / j.bbabio.2008.03.006. ISSN 0005-2728. PMID 18396149.
- ^ Годри, Майкл Дж .; Ястрох, Мартин; Треберг, Джейсон Р .; Хофрайтер, Майкл; Paijmans, Johanna L.A .; Старретт, Джеймс; Уэльс, Натан; Синьор, Энтони В .; Springer, Mark S .; Кэмпбелл, Кевин Л. (12.07.2017). «Инактивация термогенного UCP1 как историческая случайность в нескольких кладах плацентарных млекопитающих». Достижения науки. 3 (7): e16028781. Дои:10.1126 / sciadv.1602878. ЧВК 5507634. PMID 28706989.
- ^ Сайто, Сигеру; Сайто, Клэр Танака; Шингаи, Рюдзо (31.01.2008). «Адаптивная эволюция гена разъединяющего протеина 1 способствовала приобретению нового неподвижного термогенеза у предковых животных-млекопитающих». Ген. 408 (1): 37–44. Дои:10.1016 / j.gene.2007.10.018. ISSN 0378-1119. PMID 18023297.
- ^ Робинсон, Алан Дж .; Овери, Кэтрин; Кунджи, Эдмунд Р. С. (18 ноября 2008 г.). «Механизм транспорта митохондриальными переносчиками на основе анализа симметрии». Труды Национальной академии наук. 105 (46): 17766–17771. Дои:10.1073 / pnas.0809580105. ISSN 0027-8424. ЧВК 2582046. PMID 19001266.
- ^ Рикье, Даниэль (2017-03-01). «UCP1, митохондриальный разобщающий белок коричневых адипоцитов: личный вклад и историческая перспектива». Биохимия. UCP1: 40 лет и старше. 134: 3–8. Дои:10.1016 / j.biochi.2016.10.018. ISSN 0300-9084. PMID 27916641.
- ^ Николс Д.Г., Берсон В.С., Хитон Г.М. (1978). «Идентификация компонента внутренней мембраны митохондрий бурой жировой ткани, ответственного за регулирование рассеивания энергии». Experientia. Дополнение. 32: 89–93. Дои:10.1007/978-3-0348-5559-4_9. ISBN 978-3-0348-5561-7. PMID 348493.
- ^ Kozak LP, Britton JH, Kozak UC, Wells JM (сентябрь 1988 г.). «Ген митохондриального разобщающего белка. Корреляция структуры экзона с трансмембранными доменами». Журнал биологической химии. 263 (25): 12274–7. PMID 3410843.
- ^ Bouillaud F, Raimbault S, Ricquier D (декабрь 1988 г.). «Ген разобщающего белка крысы: полная последовательность, структура первичного транскрипта и эволюционные отношения между экзонами». Сообщения о биохимических и биофизических исследованиях. 157 (2): 783–92. Дои:10.1016 / S0006-291X (88) 80318-8. PMID 3202878.
- ^ Kozak LP, Anunciado-Koza R (декабрь 2008 г.). «UCP1: его участие и полезность при ожирении». Международный журнал ожирения. 32 Дополнение 7 (Дополнение 7): S32-8. Дои:10.1038 / ijo.2008.236. ЧВК 2746324. PMID 19136989.
дальнейшее чтение
- Машер, Габриэль; Келер, Мелани; Рупрехт, Энн; Крайтер, Юрген; Хинтердорфер, Питер; Поль, Елена Е. (март 2018 г.). «Ингибирование митохондриальных UCP1 и UCP3 пуриновыми нуклеотидами и фосфатом». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1860 (3): 664–672. Дои:10.1016 / j.bbamem.2017.12.001. ЧВК 6118327. PMID 29212043.
- Урбанкова, Ева; Волченко, Анна; Поль, Питер; Ежек, Петр; Поль, Елена Е. (29 августа 2003 г.). «Транспортная кинетика разобщающих белков». Журнал биологической химии. 278 (35): 32497–32500. Дои:10.1074 / jbc.M303721200. PMID 12826670.
- Ricquier D, Bouillaud F (январь 2000 г.). «Гомологи разобщающих белков: UCP1, UCP2, UCP3, StUCP и AtUCP». Биохимический журнал. 345 Pt 2 (2): 161–79. Дои:10.1042/0264-6021:3450161. ЧВК 1220743. PMID 10620491.
- Muzzin P (апрель 2002 г.). «Разъединяющие белки». Анналы д'Эндокринологии. 63 (2 Pt 1): 106–10. PMID 11994670.
- Дель Мар Гонсалес-Баррозо М., Рикье Д., Кассар-Дульсье А.М. (октябрь 2000 г.). «Ген человеческого разобщающего белка-1 (UCP1): современное состояние и перспективы исследований ожирения». Обзоры ожирения. 1 (2): 61–72. Дои:10.1046 / j.1467-789x.2000.00009.x. PMID 12119988. S2CID 30231289.
- Cassard AM, Bouillaud F, Mattei MG, Hentz E, Raimbault S, Thomas M, Ricquier D (июль 1990 г.). «Ген белка разобщения человека: структура, сравнение с геном крысы и отнесение к длинному плечу хромосомы 4». Журнал клеточной биохимии. 43 (3): 255–64. Дои:10.1002 / jcb.240430306. PMID 2380264. S2CID 31128860.
- Bouillaud F, Villarroya F, Hentz E, Raimbault S, Cassard AM, Ricquier D (июль 1988 г.). «Обнаружение мРНК несвязанного белка коричневой жировой ткани у взрослых пациентов с помощью геномного зонда человека». Клиническая наука. 75 (1): 21–7. Дои:10.1042 / cs0750021. PMID 3165741.
- Oppert JM, Vohl MC, Chagnon M, Dionne FT, Cassard-Doulcier AM, Ricquier D, Pérusse L, Bouchard C (август 1994). «Полиморфизм ДНК в гене разобщающего белка (UCP) и жировой ткани человеческого тела». Международный журнал ожирения и связанных с ним метаболических нарушений. 18 (8): 526–31. PMID 7951471.
- Клеман К., Руис Дж., Кассар-Дульсье А.М., Буйо Ф., Рикье Д., Басдеван А., Ги-Гран Б., Фрогель П. (декабрь 1996 г.). «Дополнительный эффект варианта A -> G (-3826) гена разобщающего белка и мутации Trp64Arg гена бета-3-адренорецептора на увеличение веса при патологическом ожирении». Международный журнал ожирения и связанных с ним метаболических нарушений. 20 (12): 1062–6. PMID 8968850.
- Schleiff E, Shore GC, Goping IS (март 1997 г.). «Человеческий митохондриальный импортный рецептор, Tom20p. Использование глутатиона для выявления специфических взаимодействий между Tom20-глутатион S-трансферазой и митохондриальными белками-предшественниками». Письма FEBS. 404 (2–3): 314–8. Дои:10.1016 / S0014-5793 (97) 00145-2. PMID 9119086. S2CID 29177508.
- Урхаммер С.А., Фридберг М., Соренсен Т.И., Эхвальд С.М., Андерсен Т., Тибьярг-Хансен А., Клаузен Дж.О., Педерсен О. (декабрь 1997 г.). «Исследования генетической изменчивости гена разобщающего белка 1 у субъектов европеоидной расы с ювенильным ожирением». Журнал клинической эндокринологии и метаболизма. 82 (12): 4069–74. Дои:10.1210 / jc.82.12.4069. PMID 9398715.
- Jezek P, Urbánková E (январь 2000 г.). «Специфическая последовательность мотивов митохондриальных разобщающих белков». IUBMB Life. 49 (1): 63–70. Дои:10.1080/713803586. PMID 10772343. S2CID 8541209.
- Мори Х., Окадзава Х., Ивамото К., Маэда Э., Хаширамото М., Касуга М. (март 2001 г.). «Полиморфизм в 5'-нетранслируемой области и вариант Met229 -> Leu в экзоне 5 гена UCP1 человека связаны с предрасположенностью к сахарному диабету II типа». Диабетология. 44 (3): 373–6. Дои:10.1007 / s001250051629. PMID 11317671.
- Ниббелинк М., Мулен К., Арно Э, Дюваль С., Пенико Л., Кастейла Л. (декабрь 2001 г.). «Бурый жир UCP1 специфически экспрессируется в продольных гладкомышечных клетках матки». Журнал биологической химии. 276 (50): 47291–5. Дои:10.1074 / jbc.M105658200. PMID 11572862.
- Echtay KS, Roussel D, St-Pierre J, Jekabsons MB, Cadenas S, Stuart JA, Harper JA, Roebuck SJ, Morrison A, Pickering S, Clapham JC, Brand MD (январь 2002 г.). «Супероксид активирует митохондриальные разобщающие белки». Природа. 415 (6867): 96–9. Дои:10.1038 / 415096a. PMID 11780125. S2CID 4349744.
- Руссе С., дель Мар Гонсалес-Баррозо М., Желли С., Пекер С., Буйо Ф., Рикье Д., Кассар-Дульсье А.М. (май 2002 г.). «Новый полиморфный сайт, расположенный в гене UCP1 человека, контролирует in vitro связывание CREB-подобного фактора». Международный журнал ожирения и связанных с ним метаболических нарушений. 26 (5): 735–8. Дои:10.1038 / sj.ijo.0801973. PMID 12032762.
- Rim JS, Kozak LP (сентябрь 2002 г.). «Регуляторные мотивы для CREB-связывающего белка и факторов транскрипции Nfe2l2 в вышестоящем энхансере гена митохондриального разобщающего белка 1». Журнал биологической химии. 277 (37): 34589–600. Дои:10.1074 / jbc.M108866200. PMID 12084707.
- Kieć-Wilk B, Wybrańska I, Malczewska-Malec M, Leszczyńska-Gołabek L, Partyka L, Niedbał S, Jabrocka A, Dembińska-Kieć A (сентябрь 2002 г.). «Корреляция полиморфизма -3826A> G в промоторе гена разобщающего белка 1 с ожирением и метаболическими нарушениями в семьях с ожирением из южной Польши». Журнал физиологии и фармакологии. 53 (3): 477–90. PMID 12375583.
внешняя ссылка
- Таблетки с водорослями от ожирения надежда (BBC - Термогенин упоминается как часть процесса)
- термогенин в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)