Филогенетическая номенклатура - Phylogenetic nomenclature
Филогенетическая номенклатура, часто называют кладистическая номенклатура, это метод номенклатура за таксоны в биологии, которая использует филогенетический определения названий таксонов, как описано ниже. Это контрастирует с традиционный подход, в котором названия таксонов определены тип, который может быть экземпляром или таксоном низшего классифицировать, и описание словами.[1] Филогенетическая номенклатура в настоящее время не регулируется, но Международный кодекс филогенетической номенклатуры (Филокод) предназначен для регулирования после его ратификации.
Определения
Филогенетическая номенклатура связывает имена с клады, группы, состоящие из предка и всех его потомков. Эти группы можно эквивалентно назвать монофилетический. Есть несколько разные способы указания предка, которые обсуждаются ниже. После того, как предок указан, значение имени фиксируется: предок и все организмы, являющиеся его потомками, включаются в названный таксон. Перечисление всех этих организмов (т.е. предоставление полного ограничение ) требует знания полного филогенетического дерева. На практике есть только один или несколько гипотезы что касается правильного дерева. Различные гипотезы приводят к тому, что разные организмы считаются включенными в названный таксон, но не влияют на то, к каким организмам это название действительно относится. В этом смысле название не зависит от пересмотра теории.[нужна цитата ]
Филогенетические определения названий клады
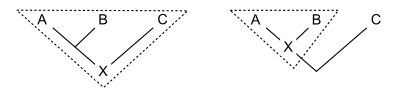
Филогенетическая номенклатура связывает имена с клады, группы, состоящие исключительно из предка и всех его потомков. Поэтому все, что нужно для определения клады, - это указать предка. Есть несколько способов сделать это. Обычно предок обозначается его отношением к двум или более спецификаторы (виды, экземпляры или признаки), которые явно упоминаются. На схеме показаны три распространенных способа сделать это. Для ранее определенных клад A, B и C кладу X можно определить как:
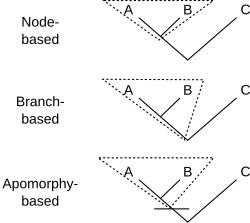
- А определение на основе узлов можно было бы прочитать: " последний общий предок A и B, и всех потомков этого предка ". Таким образом, вся линия ниже точки соединения A и B нет принадлежат к кладе, к которому относится имя с этим определением.
- Пример: зауропод динозавры состоят из последнего общего предка Вулканодон (А) и Апатозавр (В)[2] и все потомки этого предка. Этот предок был первым зауроподом. C может включать других динозавров, таких как Стегозавр.
- А определение на основе ветвей, часто называемый основанное на основе определение, можно было бы прочитать: "the первый предок A, который не является также предком C, и все потомки этого предка ". Таким образом, вся линия ниже соединения A и B (кроме самой нижней точки) делает принадлежат к кладе, к которому относится имя с этим определением.
- Пример: грызуны состоят из первых предков домовая мышь (A) который также не является предком восточный кролик (C) вместе со всеми потомками этого предка. Здесь предком является самый первый грызун. B - какой-то другой потомок, возможно, рыжая белка.
- An определение на основе апоморфии можно прочитать: «первый предок A, обладающий признаком M, который унаследован A, и все потомки этого предка». На диаграмме M развивается на пересечении горизонтальной линии с деревом. Таким образом, клада, к которой относится имя с этим определением, содержит ту часть строки, которая находится ниже последнего общего предка A и B, которая соответствует предкам, обладающим апоморфией M. Нижняя часть строки исключается. Не обязательно, чтобы B имел признак M; возможно, он исчез в линии, ведущей к B.
- Пример: четвероногие состоят из первого предка человека (A), от которого люди унаследовали конечности с пальцами рук или ног (M), и всех потомков этого предка. Эти потомки включают змеи (B), у которых нет конечностей.
Несколько других альтернатив представлены в Филокод,[3] (видеть ниже ), хотя здесь нет попытки быть исчерпывающим.
Филогенетическая номенклатура позволяет использовать не только наследственные связи, но и свойство быть сохранившийся. Один из многих способов указать Neornithes (современные птицы), например, это:
- Неорниты - это последний общий предок сохранившийся члены самой обширной клады, содержащей какаду Cacatua galerita но не динозавр Стегозавр арматус а также все потомки этого предка.
Neornithes - это корона клада, клада, для которой последний общий предок ее существующих членов также является последним общим предком все его члены.
Имена узлов
- Корона узел: Самый недавний общий предок выбранных видов интересующей клады.
- Стволовой узел: Самый недавний общий предок интересующей клады. и его сестра клада
Определения названий парафилетических и полифилетических таксонов, основанные на происхождении
в Филокод, только клады могут получить «филогенетическое определение», и это ограничение соблюдается в данной статье. Однако также возможно создать определения для названий других групп, которые являются филогенетическими в том смысле, что они используют только родственные связи, привязанные к видам или экземплярам.[4] Например, если предположить, что Mammalia и Aves (птицы) определены таким образом, Reptilia может быть определена как «самый недавний общий предок Mammalia и Aves и всех их потомков, кроме Mammalia и Aves». Это пример парафилетический группа, клада минус одна или несколько подчиненных клад. Имена полифилетический группы, характеризующиеся чертой, которая развивалась конвергентно в двух или более подгруппах аналогичным образом можно определить как сумму нескольких клад.[4]
Ранги
Под традиционным номенклатурные коды, такой как Международный кодекс зоологической номенклатуры и Международный кодекс номенклатуры водорослей, грибов и растений, таксоны, которые явно не связаны с классифицировать не могут быть официально названы, потому что применение названия к таксону основано как на типе, так и на ранге. Требование к рангу - главное различие между традиционной и филогенетической номенклатурой. Это имеет несколько последствий: ограничивает количество вложенных уровней, на которых могут применяться имена; это приводит к изменению окончаний имен, если группа изменила свой ранг, даже если у нее точно такие же члены (т. е. одинаковые ограничение ); и это логически несовместимо с тем, чтобы все таксоны были монофилетическими.
Особенно в последние десятилетия (из-за достижений в филогенетика ) таксономисты назвали множество «вложенных» таксонов (то есть таксонов, которые содержатся внутри других таксонов). Никакая система номенклатуры не пытается назвать каждую кладу; это было бы особенно сложно в традиционной номенклатуре, поскольку каждому названному таксону должен быть присвоен более низкий ранг, чем любому названному таксону, в который он вложен, поэтому количество названий, которые могут быть присвоены во вложенном наборе таксонов, не может быть больше, чем количество общепризнанных рангов. Gauthier et al. (1988)[5] предположил, что если Reptilia присваивается ее традиционный ранг класса, то филогенетическая классификация должна присвоить ранг рода Aves.[6] В такой классификации все ~ 12 000 известных видов современных и вымерших птиц должны быть включены в этот род.
Были предложены различные решения с сохранением номенклатурных кодов на основе рангов. Паттерсон и Розен (1977)[7] предложили девять новых рангов между семейством и суперсемейством, чтобы иметь возможность классифицировать кладу сельди, а McKenna and Bell (1997)[8] представил большое количество новых рангов, чтобы справиться с разнообразием Mammalia; они не получили широкого распространения. В ботанике Группа филогении покрытосеменных растений, ответственный за наиболее широко используемую в настоящее время классификацию цветущие растения, выбрал другой подход. Они сохранили традиционные звания семьи и порядка, считая их ценными для обучения и изучения взаимоотношений между таксонами, но также ввели именные клады без формальных званий.[9]
В действующих кодексах также есть правила, согласно которым названия должны иметь определенные окончания в зависимости от ранга таксонов, к которым они применяются. Если группа имеет разный ранг в разных классификациях, ее название должно иметь другой суффикс. Ерешефский (1997: 512)[6] привел пример. Он отметил, что Симпсон в 1963 году и Уайли в 1981 году согласились, что одна и та же группа родов, в которую входил род Гомо, следует поместить вместе в таксон. Симпсон относился к этому таксону как к семейству и поэтому дал ему название «Hominidae»: «Homin-» от «Homo» и «-idae» как семейство, оканчивающееся по зоологическому коду. Уайли считал, что это ранг племени, и поэтому дал ему название «Гоминини», «-ini» - окончание племени. Племя Уайли Hominini составляло лишь часть семьи, которую он называл «Hominidae». Таким образом, в соответствии с зоологическим кодексом двум группам с точно таким же охватом были даны разные названия (Гоминиды Симпсона и Гоминини Уайли), а две группы с одинаковым названием имели разные границы (Гоминиды Симпсона и Гоминиды Вили).
В филогенетической номенклатуре ранги не имеют отношения к написанию названий таксонов (см., Например, Gauthier (1994)[10] и Филокод ). Однако ранги не совсем запрещены в филогенетической номенклатуре. Они просто отделены от номенклатуры: они не влияют на то, какие названия можно использовать, какие таксоны с какими названиями связаны и какие названия могут относиться к вложенным таксонам.[11][12][13]
Принципы традиционной ранговой номенклатуры логически несовместимы с тем, что все таксоны строго монофилетичны.[11][14] Каждый организм должен принадлежать род, например, для каждого общего предка млекопитающих и птиц должен быть род. Чтобы такой род был монофилетическим, он должен включать в себя как учебный класс Mammalia и класс Aves. Однако в ранговой номенклатуре классы должны включать роды, а не наоборот.
Философия
Конфликт между филогенетической и традиционной номенклатурой отражает разные взгляды на метафизика таксонов. Для сторонников филогенетической номенклатуры таксон - это индивидуум, сущность, которая со временем приобретает и теряет атрибуты.[15] Подобно тому, как человек не становится кем-то другим, когда его или ее свойства изменяются в результате взросления, старости или более радикальных изменений, таких как амнезия, потеря конечности или изменение пола, так и таксон остается тем же существом, какие бы характеристики ни были приобретены. или потеряли.[16]
Для любого индивидуума должно быть что-то, что связывает его временные стадии, в силу чего он остается тем же самым. Для человека пространственно-временная непрерывность тела обеспечивает соответствующую связь; От младенчества до старости тело следует непрерывным путем в мире, и именно этот путь, а не какие-либо индивидуальные особенности, связывает ребенка и восьмидесятилетнего ребенка.[17] Для таксона, если характеристики не имеют отношения к делу, девонский период связывают только родственные связи. Риниогнатха хирсти с современным бабочка монарх как представители таксона Insecta, разделенные 400 миллионами лет назад.[16]
Однако если предков достаточно для непрерывности таксона, тогда все потомки члена таксона также будут включены в таксон, так что все истинные таксоны являются монофилетическими; названия парафилетических групп не заслуживают официального признания. В качестве "Пеликозаврия "относится к парафилетической группе, которая включает некоторых пермских четвероногих, но не их нынешних потомков, это не может быть признано действительным названием таксона.
С другой стороны, для приверженцев традиционной номенклатуры таксоны - это наборы или классы.[15] В отличие от индивидуумов, они состоят из сходства, характеристик, общих для их членов.[18] Монофилетические группы особенно заслуживают внимания и наименования, прежде всего потому, что они часто имеют общие свойства, представляющие интерес. Поскольку многие парафилетические группы также обладают такими свойствами, плезиоморфии в их случае предоставление им имен также способствует продуктивным исследованиям. Некоторые ученые решительно отстаивают такое именование; в письме к редакции журнала 2005 г. Таксон, 150 биологов со всего мира объединились в защиту парафилетических таксонов.[19] Они указывали, что для Дарвина эволюция связана с происхождением. и модификация, а не просто спуск. Таксоны для них - это совокупность организмов, объединенных сходством; когда сходство слишком слабое, потомки не входят во все таксоны своих предков.
История
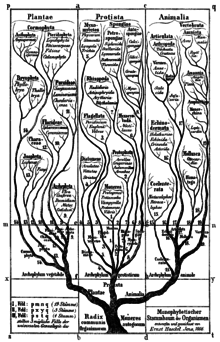
Филогенетическая номенклатура является результатом общего признания ветвления в ходе эволюции, представленной на диаграммах Жан-Батист Ламарк и более поздние писатели любят Чарльз Дарвин и Эрнст Геккель.[21] В 1866 году Геккель впервые построил единое дерево всего живого и сразу же приступил к преобразованию его в классификацию. Эта классификация была ранговой, как это было обычно в то время, но не содержала таксонов, которые Геккель считал полифилетический. В нем Геккель ввел звание филюм что несет в себе коннотацию монофилия в его названии (буквально означает «стебель»).
С тех пор ведутся дискуссии о том, каким образом и в какой степени филогения жизни должна использоваться в качестве основы для ее классификации, причем взгляды простираются от «числовой таксономии» (фенетика ) над "эволюционная таксономия «(градастика) к« филогенетической систематике ». Начиная с 1960-х годов, время от времени предлагались бесранговые классификации, но в целом принципы ранговой номенклатуры использовались всеми тремя философскими школами.
Однако большинство основных принципов филогенетической номенклатуры (отсутствие обязательных рангов и что-то близкое к филогенетическим определениям) можно проследить до 1916 года, когда Эдвин Гудрич[22] интерпретировал имя Sauropsida, возведенный 40 годами ранее Т. Хаксли, чтобы включить птиц (Авес ) а также в составе Рептилии, и придумал новое название Теропсида включить млекопитающих, а также другую часть Reptilia. Гудрич не присваивал им ранги и относился к ним точно так же, как если бы у них были филогенетические определения, не используя ни содержания, ни диагностических признаков, чтобы решить, должно ли данное животное принадлежать к Theropsida, Sauropsida или чему-то еще, после того, как его филогенетическое положение было согласовано. Гудрич также высказал мнение, что от названия Reptilia следует отказаться, как только филогения рептилий станет более известной.
Принцип, что только клады Должен быть официально назван стал популярным в определенных кругах во второй половине 20 века. Он распространился вместе с методами обнаружения клад (кладистика ) и является неотъемлемой частью филогенетической систематики (см. выше). В то же время стало очевидно, что обязательные ранги, являющиеся частью традиционных систем номенклатуры, создают проблемы. Некоторые авторы предлагали вообще отказаться от них, начиная с Вилли Хенниг отказ[23] из его более раннего предложения определить ранги как геологические возрастные классы.[24][25]
Первое использование филогенетической номенклатуры в публикации можно датировать 1986 годом.[26] Вскоре последовали теоретические статьи с изложением принципов филогенетической номенклатуры, а также дальнейшие публикации, содержащие приложения филогенетической номенклатуры (в основном к позвоночным) (см. Раздел «Литература»).
В попытке избежать раскол в сообществе биологов "Готье предложил двум членам ICZN применять формальные таксономические названия, регулируемые зоологическим кодексом, только к кладам (по крайней мере, для надвидовых таксонов) и отказаться от линнеевских рангов, но эти два члена сразу же отвергли эти идеи »(Laurin, 2008: 224).[27] Это побудило Кевина де Кейроса и ботаника Филиппа Кантино начать разработку своего собственного кодекса номенклатуры, Филокод, для регулирования филогенетической номенклатуры.
Полемика
Вилли Хенниг новаторская работа вызвала оживленные дебаты[28] об относительных достоинствах филогенетической номенклатуры по сравнению с таксономией Линнея или связанном подходе эволюционная таксономия, который продолжается до настоящего времени.[29] Некоторые дебаты, в которых были задействованы кладисты, велись с XIX века.[30] Хотя Хенниг настаивал на том, что разные схемы классификации были полезны для разных целей,[31] он отдал приоритет своему собственному, утверждая, что категории его системы обладают «индивидуальностью и реальностью» в отличие от «вневременных абстракций» основанных на морфологии классификаций.[32]
Формальные классификации, основанные на кладистических рассуждениях, как говорят, подчеркивают происхождение за счет описательных характеристик. Тем не менее, большинство таксономистов сегодня избегают парафилетических групп, когда они думают, что это возможно в рамках таксономии Линнея; полифилетические таксоны давно вышли из моды.
В Международный кодекс филогенетической номенклатуры
В ICPN, или же Филокод, представляет собой проект свода правил и рекомендаций по филогенетической номенклатуре.
- В ICPN будет только регулировать клады имена. Названия пара- или полифилетических таксонов и названия видов (которые могут быть или не быть кладами) не будут рассматриваться, по крайней мере, вначале. Это означает, что регулирование названий видов пока будет оставлено на усмотрение ранговых коды номенклатуры.
- В Принцип приоритета будут введены для имен и для определений. Отправной точкой для определения приоритета будет дата публикации ICPN.
- Определения существующих имен и новых имен вместе с их определениями должны быть опубликованы в рецензируемый работает (на дату начала или позже) и должна быть зарегистрирована в онлайн-базе данных, чтобы быть действительной.
Число сторонников повсеместного принятия Филокод все еще невелик, и неизвестно (по состоянию на ноябрь 2020 года), когда код будет реализован и насколько широко он будет применяться.
Рекомендации
- ^ Международная комиссия по зоологической номенклатуре (1999). «Глоссарий». Международный кодекс зоологической номенклатуры (4-е изд.). ISBN 978-0-85301-006-7.CS1 maint: ref = harv (связь)
- ^ Бентон, Майкл Дж. (2005). Палеонтология позвоночных. Блэквелл. п. 214. ISBN 978-0-632-05637-8.
- ^ Кантино, Филип Д. и де Кейрос, Кевин (2010). «Международный кодекс филогенетической номенклатуры, версия 4c». примечание 9.3.1.
| вклад =игнорируется (помощь)CS1 maint: ref = harv (связь). - ^ а б de Queiroz, K .; Готье, Дж. (1990). «Филогения как центральный принцип в таксономии: филогенетические определения названий таксонов». Систематическая зоология. 39 (4): 307–322. Дои:10.2307/2992353. JSTOR 2992353.CS1 maint: ref = harv (связь)
- ^ Готье, Дж., Эстес, Р. и де Кейро, К., 1988. Филогенетический анализ лепидозавроморф. Стр. 15–98 в R. Estes & G. Pregill (ред.): Филогенетические взаимоотношения семейств ящериц: очерки памяти Чарльза Л. Кэмпа. Stanford University Press. ISBN 978-0-8047-1435-8
- ^ а б Ерешефский, М. (1997). «Эволюция Линнеевской иерархии». Биология и философия. 12 (4): 493–519. Дои:10.1023 / А: 1006556627052. S2CID 83251018.CS1 maint: ref = harv (связь)
- ^ Паттерсон, К. и Розен, Д. Обзор ихтиодектиформ и других мезозойских костистых рыб, а также теория и практика классификации окаменелостей, 1977 г. Бюллетень Американского музея естественной истории 158: 81–172.
- ^ Маккенна, М.С. и Белл, С.К. 1997. Классификация млекопитающих выше видового уровня. Издательство Колумбийского университета. ISBN 0-231-11012-X
- ^ Группа филогении покрытосеменных (1998). «Порядковая классификация семейств цветковых растений». Летопись ботанического сада Миссури. 85 (4): 531–553. Дои:10.2307/2992015. JSTOR 2992015. S2CID 82134384.CS1 maint: ref = harv (связь)
- ^ Готье, Дж. А. 1994. Диверсификация амниот. Стр. 129–159 в: Д. Р. Протеро и Райнер Р. Шох (ред.): Основные особенности эволюции позвоночных. Палеонтологическое общество.
- ^ а б de Queiroz, K .; Готье, Дж. (1992). «Филогенетическая таксономия [sic ]". Анну. Rev. Ecol. Syst. 23: 449–480.CS1 maint: ref = harv (связь)
- ^ Кантино, П. Д. (2000). «Филогенетическая номенклатура: решение некоторых проблем». Таксон. 49 (1): 85–93. Дои:10.2307/1223935. JSTOR 1223935.CS1 maint: ref = harv (связь)
- ^ Bryant, H.N .; Кантино, П. Д. (2002). «Обзор критики филогенетической номенклатуры: является ли таксономическая свобода фундаментальным вопросом?». Биол. Rev. 77 (1): 39–55. Дои:10.1017 / S1464793101005802. PMID 11911373. S2CID 20518066.CS1 maint: ref = harv (связь)
- ^ Казлев, М.А. «Кладистическая и линнеевская системы - несовместимы или дополняют друг друга?». (palaeos.com). Архивировано из оригинал 10 июля 2017 г.. Получено 30 сентября, 2012.
- ^ а б Assis, L.C.S .; Бригандт, И. (2009). "Гомология: виды кластеров гомеостатических свойств в систематике и эволюции" (PDF). Эволюционная биология. 36 (2): 248–255. Дои:10.1007 / s11692-009-9054-у. S2CID 363300.CS1 maint: ref = harv (связь)[постоянная мертвая ссылка ]
- ^ а б Роу, Тимоти (1988). «Определение, диагноз и происхождение млекопитающих» (PDF). Журнал палеонтологии позвоночных. 8 (3): 241–264. Дои:10.1080/02724634.1988.10011708.CS1 maint: ref = harv (связь)
- ^ Уиггинс, Дэвид (1967). Идентичность и пространственно-временная непрерывность. Издательство Оксфордского университета. ISBN 978-0631103707.CS1 maint: ref = harv (связь)
- ^ Запись для «таксона» в: Международная комиссия по зоологической номенклатуре (1999). «Глоссарий». Международный кодекс зоологической номенклатуры (4-е изд.). ISBN 978-0-85301-006-7.CS1 maint: ref = harv (связь)
- ^ Нордал, Ингер и Стедье, Брита, координаторы (2005). «Парафилетические таксоны должны быть приняты». Таксон. 54 (1): 5–8. Дои:10.2307/25065296. JSTOR 25065296.CS1 maint: несколько имен: список авторов (связь) CS1 maint: ref = harv (связь)
- ^ Геккель, Э. Х. Ф. А. 1866. Generelle Morphologie der Organismen. Георг Реймер.
- ^ Раган, Марк А. (2009). «Деревья и сети до и после Дарвина». Биология Директ. 4 (43): 43. Дои:10.1186/1745-6150-4-43. ЧВК 2793248. PMID 19917100.CS1 maint: ref = harv (связь)
- ^ Гудрич, Э. С. (1916). «О классификации рептилий». Труды Королевского общества B. 89 (615): 261–276. Bibcode:1916RSPSB..89..261G. Дои:10.1098 / rspb.1916.0012.CS1 maint: ref = harv (связь)
- ^ Хенниг, В. 1969. Die Stammesgeschichte der Insekten. Вальдемар Крамер.
- ^ Хенниг, В. 1950. Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag.
- ^ Хенниг, В. (1965). «Филогенетическая систематика». Ежегодный обзор энтомологии. 10: 97–116. Дои:10.1146 / annurev.en.10.010165.000525.CS1 maint: ref = harv (связь)
- ^ Готье, Ж. 1986. Заурисхийская монофилия и происхождение птиц. Стр. 1–55 дюймов К. Падиан (ред.): Происхождение птиц и эволюция полета. Мемуар 8 Калифорнийской академии наук.
- ^ Лаурин, М. (2008). «Великолепная изоляция биологической номенклатуры». Zoologica Scripta. 37 (2): 223–233. Дои:10.1111 / j.1463-6409.2007.00318.x. S2CID 85020798.CS1 maint: ref = harv (связь)
- ^ Уилер, Квентин (2000). Видовые концепции и филогенетическая теория: дискуссия. Издательство Колумбийского университета. ISBN 978-0-231-10143-1.CS1 maint: ref = harv (связь)
- ^ Бентон, М. Дж. (2000). «Стебли, узлы, коронные клады и списки без рангов: Линней мертв?» (PDF). Биологические обзоры. 75 (4): 633–648. CiteSeerX 10.1.1.573.4518. Дои:10.1111 / j.1469-185X.2000.tb00055.x. PMID 11117201. S2CID 17851383. Архивировано из оригинал (PDF) на 2017-08-09. Получено 2011-08-26.CS1 maint: ref = harv (связь)
- ^ Халл, Дэвид (1988). Наука как процесс. Издательство Чикагского университета. С. 232–276. ISBN 978-0-226-36051-5.CS1 maint: ref = harv (связь)
- ^ Хенниг, Вилли (1966). Филогенетическая систематика (тр. Д. Дуайт Дэвис и Райнер Зангерл). Урбана, Иллинойс: Univ. of Illinois Press (перепечатано в 1979 и 1999 гг.). п. 9. ISBN 978-0-252-06814-0.CS1 maint: ref = harv (связь)
- ^ Хенниг 1966, п. 81 год
дальнейшее чтение
Здесь цитируется несколько публикаций, не цитируемых в ссылках. Исчерпывающий список публикаций по филогенетической номенклатуре можно найти на интернет сайт из Международное общество филогенетической номенклатуры.
- Брайант, Гарольд Н. (1994). «Комментарии к филогенетическому определению названий таксонов и соглашениям, касающимся наименования кладовых крон». Syst. Биол. 43: 124–129. Дои:10.1093 / sysbio / 43.1.124.CS1 maint: ref = harv (связь)
- Кантино, Филип Д .; Олмстед, Ричард Г. (2008). «Применение филогенетически определенных имен не требует, чтобы каждый спецификатор присутствовал на дереве». Syst. Биол. 57 (1): 157–160. Дои:10.1080/10635150701883873. PMID 18300028.CS1 maint: ref = harv (связь)
- де Кейруш, Кевин (1992). Филогенетические определения и таксономическая философия. Биол. Филос. 7:295–313.
- Готье, Жак А., Арнольд Г. Клюге и Тимоти Роу (1988). Ранняя эволюция амниот. Страницы 103–155 в Майкл Дж. Бентон (ред.): Филогения и классификация четвероногих, том 1: амфибии, рептилии, птицы. Syst. Жопа. Спец. Vol. 35A. Кларендон Пресс, Оксфорд.
- Готье, Жак, Дэвид Каннателла, Кевин де Кейроз, Арнольд Г. Клюге и Тимоти Роу (1989). Филогения четвероногих. Страницы 337–353 в Б. Фернхольм, К. Бремер и Х. Йорнвалль (ред.): Иерархия жизни. Elsevier Science B.V. (Биомедицинский отдел), Нью-Йорк.
- Гизелин, М. Т. (1984). «Определение», «характер» и другие двусмысленные термины. Syst. Zool. 33 (1): 104–110. Дои:10.2307/2413135. JSTOR 2413135.CS1 maint: ref = harv (связь)
- Кизи, Т. Майкл (2007). «Математический подход к определению названий клад с потенциальными приложениями к компьютерному хранению и обработке». Zool. Scr. 36 (6): 607–621. Дои:10.1111 / j.1463-6409.2007.00302.x. S2CID 83862527.CS1 maint: ref = harv (связь)
- Лорен, Мишель (2005). Преимущества филогенетической номенклатуры перед номенклатурой Линнея. Страницы 67–97 в А. Минелли, Дж. Орталли и Г. Санга (ред.): Имена животных. Instituto Veneto di Scienze, Lettere ed Arti; Венеция.
- Ли, Майкл С. Ю. (2005). «Выбор референтных таксонов в филогенетической номенклатуре». Zool. Scr. 34 (3): 329–331. Дои:10.1111 / j.1463-6409.2005.00196.x. S2CID 86329828.CS1 maint: ref = harv (связь)
- Роу, Тимоти (1987). «Определение и диагностика в филогенетической системе». Syst. Zool. 36 (2): 208–211. Дои:10.2307/2413270. JSTOR 2413270.CS1 maint: ref = harv (связь)
- Роу, Тимоти; Готье, Жак (1992). «Происхождение, палеонтология и определение названия Mammalia». Syst. Биол. 41 (3): 372–378. Дои:10.1093 / sysbio / 41.3.372. S2CID 86132781.CS1 maint: ref = harv (связь)
- Серено, Пол С. (1998). «Обоснование филогенетических определений в применении к таксономии динозавров более высокого уровня». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 210: 41–83. Дои:10.1127 / njgpa / 210/1998/41.CS1 maint: ref = harv (связь)
- Серено, Пол С. (1999). «Определения в филогенетической таксономии: критика и обоснование». Syst. Биол. 48 (2): 329–351. Дои:10.1080/106351599260328. PMID 12066711.CS1 maint: ref = harv (связь)
- Серено, Пол С. (2005). "Логическая основа филогенетической систематики [sic]". Syst. Биол. 54 (4): 595–619. Дои:10.1080/106351591007453. PMID 16109704.CS1 maint: ref = harv (связь)
- Тейлор, Майкл П. (2007). «Филогенетические определения в эпоху до PhyloCode; последствия для наименования клад под PhyloCode». Палеобиос. 27: 1–6.CS1 maint: ref = harv (связь)
- Уилкинсон, Марк (2006). «Определение стабильных референтных таксонов для филогенетической номенклатуры». Zool. Scr. 35: 109–112. Дои:10.1111 / j.1463-6409.2005.00213.x. S2CID 85702468.CS1 maint: ref = harv (связь)
- Wyss, A. R .; Менг, Дж. (1996). «Применение филогенетической таксономии к плохо определенным кладам кроны: определение Rodentia на основе модифицированных стеблем узлов». Syst. Биол. 45 (4): 559–568. Дои:10.1093 / sysbio / 45.4.559.CS1 maint: ref = harv (связь)
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||