Орнитолестес - Ornitholestes
| Орнитолестес | |
|---|---|
 | |
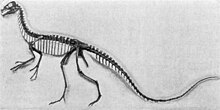
| Крепление на скелет, Королевский музей Тиррелла | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Clade: | Maniraptoromorpha |
| Подсемейство: | †Ornitholestinae Павел, 1988 |
| Род: | †Орнитолестес Осборн, 1903 |
| Типовой вид | |
| †Ornitholestes hermanni Осборн, 1903 г. | |
Орнитолестес (что означает «птичий грабитель») - небольшой теропод динозавр последнего Юрский (Brushy Basin Member из Формация Моррисон, середина Кимериджский возраст, около 154 миллионов лет назад[1]) западных Лавразия (область, которая должна была стать Северная Америка ).
Назначить свидание, Орнитолестес известен только по единичному частичному скелету с сильно раздробленным черепом, найденным на Костяной карьер возле Лук медицины, Вайоминг, в 1900 году. Его описал Генри Фэрфилд Осборн в 1903 году. Незавершенная рука позже была приписана Орнитолестес, хотя теперь кажется, что он принадлежит Таниколагрей. В тип (и единственный известный) вид О. Германни. В конкретное имя чтит Американский музей естественной истории препарат Адам Германн.
Описание

Орнитолестес был двуногим хищником.[2] Его голова была пропорционально меньше, чем у большинства других хищных динозавров, но череп был плотно сложен, с короткой мордой и крепкой нижней челюстью.[3] В орбиты (глазницы) были довольно большими, их длина составляла более 25 процентов длины черепа.[4] Нет никаких указаний на костлявое кольцо для глаз.[5]
Передние зубы Орнитолестес были несколько коническими, с уменьшенными зазубринами; задние зубы были загнутыми и более острыми, как у других динозавров-теропод.[6] Генри Фэрфилд Осборн (1903) насчитал четыре зуба в предчелюстная кость, из которых передний зуб был самым большим в верхней челюсти.[7] В отличие, Грегори С. Пол (1988) изобразил череп только с тремя оставшимися предчелюстными зубами, намного меньшими, чем те, что проиллюстрированы Осборном.[8] Каждый верхняя челюсть (основная зубная кость верхней челюсти) содержала десять зубов, каждый зубной (зубоносная кость нижней челюсти) содержала двенадцать зубов.[9] Зубные ряды Орнитолестес были короткими, при этом зубной (нижний) ряд был даже короче верхнечелюстного (верхнего) ряда,[10] несмотря на то, что сама зубная кость была исключительно длинной сзади, достигая точки ниже середины глазницы.[11] Зубы не доходили до орбит, и ни один из зубных рядов не занимал больше одной трети черепа.[12]

Область сломанной кости рядом с наружной ноздрей, кажется, выпирает вверх, что побудило Грегори С. Пола предположить, что Хищные динозавры мира (1988), что Орнитолестес носовой рог был «похож на куриный гребень».[13] Оба Oliver W.M. Раухут (2003) и Кеннет Карпентер и другие. (2005а) отверг эту интерпретацию и указал, что восходящее выступание кости произошло из-за посмертного раздавливания черепа.[14] Обновленная иллюстрация Павла Орнитолестес в его 2010 Принстонский полевой путеводитель по динозаврам больше не содержит носовой рог.[15]
Орнитолестес имел относительно короткую шею с небольшой сигмовидной (S-образной) кривой.[16] Хвост был длинным и напоминающим хлыст, составлял более половины длины тела.[17] Не все позвонки сохранились, но Осборн (1917) подсчитал, что Орнитолестес имел девять или десять шейных (шейных) позвонков, тринадцать спинных (спинных) позвонков, четыре крестцовых (бедренных) позвонка и от 39 до 44 хвостовых (хвостовых) позвонков.[18] плотник и другие. (2005а) зафиксировал, что образец содержал пять крестцовых позвонков.[19] Орнитолестес был короткотелым тероподом, и это отражалось в коротких размерах шейных и спинных позвонков спереди назад.[20]

Передние конечности Орнитолестес были относительно длинными, чуть менее двух третей длины задних ног.[21] В плечевая кость (кость плеча) была мощной и несколько длиннее, чем радиус и локтевая кость (кости предплечья).[22] И плечевая кость, и лучевая кость имели прямой стержень.[23] Когти на I и II пальцах руки были примерно одинакового размера.[24] Хотя третий ноготь руки (кость копыта) не сохранился, экстраполяция от ближайших родственников Орнитолестес указывает, что он, вероятно, был короче первых двух.[24]
Орнитолестес часто изображается как быстрый, длинноногий теропод, но кости его нижних конечностей были довольно короткими.[25] Осборн (1917) подсчитал, что пропавшие без вести большеберцовая кость (большеберцовая кость) составляла всего около 70,6% от длины бедренная кость (бедренная кость).[26] В плюсневые кости (кости стопы) расположены близко друг к другу, но не срослись.[27] Как типично для теропод, лапы были трехпалые (с тремя когтистыми несущими пальцами).[28] Джон Х. Остром (1969) отметил, что коготь на пальце II (самый внутренний палец) был больше, чем на пальцах III и IV, и предположил, что на этом пальце мог быть видоизмененный серповидный коготь, похожий на коготь когтя. Дейноних.[29] Однако, как отметили Остром (1969) и Пол (1988), плохая сохранность второго пальца затрудняет подтверждение этой гипотезы.[30]

В своем описании 1903 года Осборн писал, что длина Орнитолестес вдоль «черепа и позвоночника в восстановленном виде» составлял 2,22 м (7,28 фута).[31] Однако эта реконструкция была неточной, поскольку частично основывалась на Отниэль Чарльз Марш восстановление базальный завроподоморф Анхизавр, и шея, и туловище были слишком удлиненными.[32] Дэвид Норман (1985) и Джон Фостер (2007) оценили, что Орнитолестес был около 2 м (6,6 футов) в длину.[33] Грегори С. Пол 1988 Хищные динозавры мира перечислил длину Орнитолестес примерно 2,08 м (6,82 фута).[34]
Пол (1988) и Фостер (2007) оценили, что Орнитолестес весил 12,6 кг (27,8 фунта).[35] Джон А. Лонг и Питер Схоутен (2008) предложил немного более высокую цифру - 15 кг (33 фунта).[36]
Открытие и наименование

Орнитолестес был первым тероподом, обнаруженным в 1900-х годах.[37] В голотип скелет (AMNH 619) был раскопан в июле 1900 г. в г. Костяной карьер в Вайоминг по Американский музей естественной истории экспедиция Питера К. Кайзена, Пола Миллера и Фредерик Брюстер Лумис.[31][38] Он представляет собой частичный скелет с черепом, включающим многочисленные элементы позвоночника, передних конечностей, таза и задних конечностей. Генри Фэрфилд Осборн назвал и научно описал этот экземпляр в 1903 году.[39] В род имя Орнитолестес, первоначально предложенный Теодор Гилл, означает «птичий грабитель» и происходит от Греческий ὄρνις/Орнис, орнитос ("птица") и λῃστήσ/Lestes («грабитель»).[40] В разновидность имя (О. Германни) чествует Адама Германна, главного подготовителя музея, руководившего реставрацией и установкой скелета.[31]
Неполная рука (AMNH 587) была назначена Орнитолестес Осборном в его описании рода в 1903 году.[41] Однако, как заметил Грегори С. Пол (Gregory S. Paul, 1988), плохая сохранность соответствующих элементов в типовом образце сделала эту ассоциацию «предварительной».[42] В 2005 году Кеннет Карпентер и другие. описал нового маленького теропод, Таниколагрей, чей скелет был найден в карьере Костяной Хижины всего в нескольких сотнях ярдов от AMNH 587.[43] Поскольку AMNH 587 был практически идентичен сохранившейся руке Таниколагрей типовой образец, теперь считается, что он принадлежит этому динозавру, а не Орнитолестес.[44] После этого переназначения Фил Сентер (2006) отметил, что «наши знания о Орнитолестес теперь можно нарисовать только с голотипа ".[45] Джон Фостер (2007) сообщил, что некоторые фрагменты из Карьер Dry Mesa может принадлежать Орнитолестес,[46] хотя они еще не описаны.
В 1920 г. Чарльз Уитни Гилмор пришли к выводу, что Орнитолестес был идентичен Целурус;[47] в 1934 г. Оливер Перри Хэй признал разницу только на уровне видов, назвав Coelurus hermanni,[48] но в 1980 году Джон Остром возродил род.[49]
Классификация
В инфраотряд Целурозаврия, изобретенный в 1914 г. Фридрих фон Хюене, традиционно был таксономическая корзина для мусора в которую были помещены все мелкие теропод.[50] Орнитолестеспоэтому из-за своего небольшого размера обычно классифицируется как целурозавр.[51] В 1986 г. Жак Готье переопределил этот и несколько других палеонтологических терминов более строго, основываясь на кладистический методы. Столбняк был определен как современные птицы и все тероподы более тесно связаны с современными птицами, чем с кератозавры, в то время как целурозаврия теперь включала всех представителей столбняка, более тесно связанных с современными птицами, чем с карнозавры.[52] В 1988 г. Грегори С. Пол Предполагается, что Орнитолестес был очень похож по строению черепа на Процератозавр, среднеюрский теропод из Англии.[5] Он поместил эти два рода вместе в Ornitholestinae - новое подсемейство под Аллозавриды - и предположил, что они были более тесно связаны с гораздо более крупными Аллозавр чем другим мелким тероподам.[53] Однако классификация Орнитолестес и Процератозавр поскольку родственники аллозавров оказались несостоятельными (последний с тех пор оказался тираннозавроид ), и Пол в конце концов отказался от нее.[54] Все опубликованные кладистические анализы показали Орнитолестес быть целурозавром по определению Готье. Некоторые анализы подтвердили гипотезу о том, что это самый примитивный член группы. Манираптора, хотя более тщательный анализ показал, что он более примитивен, чем Maniraptoriformes, и, возможно, близкий родственник "compsognathid" Юравенатор Старкий.[55]
Следующее генеалогическое древо иллюстрирует синтез взаимоотношений основных групп целурозавров на основе различных исследований, проведенных в 2010-х годах.[56]
| Целурозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||


Палеобиология
В исследовании 2001 года, проведенном Брюсом Ротшильдом и другими палеонтологами, 20-футовые кости относились к Орнитолестес были исследованы на наличие признаков стресс-перелом, но ничего не было найдено.[57]
Функция передних конечностей
В биомеханическом исследовании, проведенном Филом Сентером (2006), использовались шарнирные слепки Орнитолестес наберите правую переднюю конечность образца, чтобы определить диапазон ее движений.[58] Сентер обнаружил, что переднее плечо (предплечье) может свободно качаться в пределах 95 °.° ассортимент.[59] Когда согнутый (согнуты внутрь) в локтевом суставе до максимально возможной степени плечевая кость (кость плеча) и лучевая кость (кость нижней руки) образовывали угол 53 °.[59] Способность Орнитолестес сгибание предплечья под углом, значительно более острым, чем 90 °, характерно для Maniraptoriformes, но отсутствует у более примитивных теропод, таких как Целофиз и Аллозавр.[60]
Даже когда полностью расширенный (выпрямлен) в локте, предплечье не образовывало прямой угол, отставая от этого на 22 °.[59] Пронация (скручивание, чтобы ладонная сторона руки смотрела вниз) предплечья было невозможно, потому что лучевая и локтевая кости не имели поверхностей качения, что означало, что предплечье находилось в постоянном состоянии. супинация.[24]
Когда Орнитолестес согните руки в локтях, это заставит предплечья двигаться внутрь, к средней линии.[61] Возможно, он использовал эту способность, чтобы схватить добычу обеими руками одновременно.[62]
Рацион питания

Генри Фэрфилд Осборн в своем описании 1903 г. Орнитолестесотметил его большие конические передние зубы, «быструю хватательную силу» его руки и «уравновешивающую силу» его хвоста; он интерпретировал это как приспособление к добыче современных птиц.[31] Позже Осборн отверг эту гипотезу, предположив в 1917 г. Орнитолестес представили ранние стадии перехода от плотоядного к более травоядному образу жизни, но не ранее Чарльз Р. Найт нарисовал влиятельную и широко опубликованную иллюстрацию Орнитолестес погоня Археоптерикс.[63] Иллюстрация Найта и другие, заимствованные из нее, продолжали появляться в книгах о динозаврах, ориентированных на широкую публику, на протяжении всего 20 века.[51]
Дэвид Норман (1985) признал, что это «просто возможно, хотя и маловероятно». Орнитолестес могли поймать и съесть примитивных птиц.[64] Однако самые последние авторы вместо этого предложили диету из мелких наземных позвоночных. Млекопитающие, ящерицы, лягушки, саламандры, ринхоцефалы, а вылупившиеся динозавры были бы потенциальной добычей.[65] Грегори С. Пол (1988) думал, что Орнитолестес мог использовать свои конические передние зубы для ловли рыбы.[13] Норман (1985) предположил, что крепкий череп и челюсти могли позволить Орнитолестес чтобы схватить «более крупную и более активную добычу», чем другие мелкие теропод.[66] В этом ключе Дэвид Ламберт (1993) предположил, что Орнитолестес, если бы это был стайный охотник, возможно, он смог бы орнитоподы размером с полувзрослый Камптозавр."[67]
В 2007 году Джон Фостер, специалист по формации Моррисон, предположил возможность перегородка ниши между Орнитолестес и его современный Целурус, который был в том же диапазоне размеров.[68] Он предположил, что глазастые Орнитолестес мог быть специализирован для ночной образ жизни охота, пока Целурус могли сосредоточиться на тех видах добычи, которые были активны в течение дня.[68] Фостер, однако, заметил, что эта гипотеза была в значительной степени спекулятивной; отсутствие сохранившихся черепных материалов из Целурус делает невозможным проверить, были ли его глаза пропорционально меньше, чем у Орнитолестес.[69]
Пол (1988) оценил, что 12 кг (26 фунтов) эндотермический Орнитолестес будет иметь ежедневную диетическую потребность около 700 г (1,54 фунта) мяса.[70]
Перья
Орнитолог Перси Лоу предположил в 1944 г., что Орнитолестес мог иметь перья.[71] Однако эта интерпретация в основном игнорировалась более полувека; большинство реконструкций динозавров теропод, в том числе Орнитолестес, изображал их с чешуйчатой кожей рептилий. Одним из немногих исключений из этого правила был Грегори С. Пол. Хищные динозавры мира (1988).[72] Роберт Т. Баккер с Ереси динозавров (1986) точно предсказали присутствие перьев у дромеозавридов и утверждали, что все динозавры были эндотермическими, но не использовали перья в своей иллюстрации Орнитолестес.[73]
В 1996 году примитивный целурозавр Синозауроптерикс был обнаружен в Китае. Хорошо сохранившаяся окаменелость показала, что этот динозавр был покрыт рудиментарными меховыми перьями. В результате этого и других подобных открытий большинство палеонтологов теперь считают вероятным, что все целурозавры имели те или иные изолирующие перья или произошли от предков, которые имели их.[74] Основываясь на его филогенетическом положении, Джон Фостер (2007) сделал вывод, что перья Орнитолестес вероятно, был бы более примитивным, чем у птиц; они, вероятно, «покрыли бы все тело, за исключением ног, короткой шубкой, возможно, с более длинными перьями, выстилающими верхнюю часть черепа или шеи и задний край предплечий».[75] Если предположить, что они присутствовали, эти перья могли быть использованы для изоляции, а также, возможно, для вынашивающие яйца.[75]
Рекомендации
- ^ Тернер К.Э. и Петерсон Ф. (1999). «Биостратиграфия динозавров в верхнеюрской формации Моррисона в Западных внутренних районах, США». Стр. 77–114 в Gillette, D.D. (ред.), Палеонтология позвоночных в Юте. Разное Публикация Геологической службы Юты 99-1.
- ^ Ламберт (1993), стр. 79
- ^ Пол (1988а), п. 306; Норман (1985), стр. 42; Норман (1990), стр. 293
- ^ Осборн (1903), стр. 460; Чуре (1998), стр. 238
- ^ а б Пол (1988б), п. 6
- ^ Пол (1988б), п. 3; Норман (1990), стр. 293
- ^ Осборн (1903), стр. 460; Осборн (1917), стр. 734
- ^ Пол (1988б), п. 4; Норман (1990), стр. 293
- ^ Осборн (1903), стр. 460
- ^ Пол (1988б), п. 3; Норман (1990), стр. 292
- ^ Криштиану дал Сассо и Симоне Магануко, 2011 год, Сципионикс самнитический (Theropoda: Compsognathidae) из нижнего мела Италии - остеология, онтогенетическая оценка, филогения, анатомия мягких тканей, тафономия и палеобиология, Memorie della Società Italiana de Scienze Naturali e del Museo Civico di Storia Naturale di Milano XXXVII(I): 1-281
- ^ Осборн (1903), стр. 460; Норман (1990), стр. 293
- ^ а б Пол (1988а), п. 303
- ^ Раухут (2003), стр. 27; плотник и другие. (2005а), п. 53
- ^ Пол (2010), стр. 123
- ^ плотник и другие. (2005а), стр.49, 54
- ^ Осборн (1917), стр. 734; Ламберт (1993), стр. 78
- ^ Осборн (1917), стр. 735–736
- ^ плотник и другие. (2005а), п. 58
- ^ плотник и другие. (2005а), п. 49, 69
- ^ Остром (1969), стр. 146; Пол (2002), стр. 236
- ^ Осборн (1917), стр. 738; плотник и другие. (2005б), п. 45
- ^ плотник и другие. (2005а), п. 60, 62
- ^ а б c Сентер (2006), стр. 1030
- ^ Пол (1988а), п. 306
- ^ Осборн (1917), стр. 737
- ^ плотник и другие. (2005а), п. 67
- ^ Осборн (1903), стр. 463
- ^ Остром (1969), стр. 161
- ^ Остром (1969), стр. 161; Пол (1988б), п. 6
- ^ а б c d Осборн (1903), стр. 459
- ^ Осборн (1917), стр. 735; Пол (1988б), п. 3
- ^ Норман (1985), стр. 39; Фостер (2007), стр. 178
- ^ Пол (1988а), п. 305
- ^ Пол (1988а), п. 305; Фостер (2007), стр. 178
- ^ Лонг и Схоутен (2008), стр. 73
- ^ Фастовский и Вайшампель (2005), стр. 291
- ^ Пол Д. Бринкман, 2010 г., Вторая волна динозавров юрского периода: музеи и палеонтология в Америке на рубеже ХХ века. University of Chicago Press, 345 стр., ISBN 978-0-226-07472-6
- ^ Осборн (1903), стр. 459–464
- ^ Осборн (1903), стр. 459; Осборн (1917), стр. 733
- ^ Осборн (1903), стр. 459; Осборн (1917), стр. 736
- ^ Пол (1988б), п. 3
- ^ плотник и другие. (2005б), п. 23, 39
- ^ плотник и другие. (2005б), п. 39
- ^ Сентер (2006), стр. 1029
- ^ Фостер (2007), стр. 78, 179
- ^ Гилмор, C.W., 1920, «Остеология хищных динозавров в Национальном музее США, с особым упором на роды Antrodemus (Allosaurus) и Ceratosaurus», Бюллетень Национального музея США CX: 1-154
- ^ Хэй, О.П., 1930, Вторая библиография и каталог ископаемых позвоночных Северной Америки. Институт Карнеги Вашингтона. 390 (II): 1-1074
- ^ Остром (1980)
- ^ Пол (1988а), п. 188–189; Норман (1990), стр. 280
- ^ а б Glut (1997), стр. 645
- ^ Holtz и другие. (2004), стр. 71
- ^ Пол (1988а), п. 302; Пол (1988б), п. 3, 6
- ^ Пол (2010), стр. 124
- ^ Годфруа, Паскаль; Кау, Андреа; Ху, Дун-Ю; Эскилье, Франсуа; Ву, Венхао; Дайк, Гарет (2013). «Юрский птичий динозавр из Китая решает раннюю филогенетическую историю птиц». Природа. 498 (7454): 359–362. Bibcode:2013Натура.498..359G. Дои:10.1038 / природа12168. PMID 23719374.
- ^ Хендрикс, К., Хартман, С.А., и Матеус, О. (2015). Обзор открытий и классификации теропод, не относящихся к птицам. Журнал PalArch по палеонтологии позвоночных, 12(1): 1-73.
- ^ Ротшильд, Б., Танке, Д.Х., и Форд, Т.Л., 2001, Стрессовые переломы теропод и разрывы сухожилий как ключ к активности: В: Мезозойская жизнь позвоночных, под редакцией Танке, Д.Х., и Карпентер, К., Indiana University Press, п. 331-336.
- ^ Сентер (2006), стр. 1029–1030
- ^ а б c Сентер (2006), стр. 1030–1031
- ^ Сентер (2006), стр. 1032
- ^ Сентер (2006), стр. 1030–1032
- ^ Сентер (2006), стр. 1031
- ^ Осборн (1917), стр. 733–734; Glut (1997), стр. 645
- ^ Норман (1985), стр. 39
- ^ Норман (1985), стр. 39; Пол (1988), стр. 306; Фостер (2007), стр. 179; Лонг и Схоутен (2008), стр. 73
- ^ Норман (1985), стр. 42
- ^ Ламберт (1993), стр. 78
- ^ а б Фостер (2007), стр. 180
- ^ Фостер (2007), стр. 180–181
- ^ Пол (1988а), п. 404
- ^ Витмер (1992), стр. 444
- ^ Пол (1988а), п. 122–123, 211
- ^ Баккер (1986), стр. 99, 310
- ^ Чаттерджи и Темплин (2004), стр. 277; Фастовский и Вайшампель (2005), стр. 329–330; Карри (2005), стр. 368
- ^ а б Фостер (2007), стр. 179
Источники
- Осборн, Генри Фэрфилд (1903). "Ornitholestes hermanni, новый динозавр-компсогнатоид из верхней юры ». Бюллетень Американского музея естественной истории. 19 (12): 459–464. HDL:2246/1502.
- Осборн, Генри Фэрфилд (1917). "Скелетные адаптации Орнитолестес, Струтиомим, Тиранозавр". Бюллетень Американского музея естественной истории. 35 (43): 733–771. HDL:2246/1334.
- Остром, Джон Х. (1969). «Остеология Deinonychus antirrhopus, необычный теропод из нижнего мела Монтаны ». Бюллетень музея естественной истории Пибоди. 30: 1–165.
- Остром, Джон Х. (1980). "Целурус и Орнитолестес: Они такие же? ». В Джейкобс, Луи Л. (ред.). Аспекты истории позвоночных: очерки в честь Эдвина Харриса Колберта. Музей прессы Северной Аризоны. С. 245–256. ISBN 978-0-89734-052-6.
- Норман, Дэвид Б. (1985). «Целурозавры». Иллюстрированная энциклопедия динозавров. Саламандра Букс Лтд., Стр. 38–43. ISBN 978-0-517-46890-6.
- Пол, Грегори С. (1988а). «Орнитолестиане и аллозавры». Хищные динозавры мира. Саймон и Шустер. стр.302–318. ISBN 978-0-671-61946-6. Проверить значения даты в:
| год =(Помогите) - Пол, Грегори С. (1988б). «Маленькие хищные динозавры среднего мезозоя: рогатые тероподы Моррисона и Грейт Уолита -Орнитолестес и Процератозавр- и серповидно-когтистые тероподы рек Кловерли, Джадохта и Юдифь -Дейноних, Велоцираптор и Saurornitholestes". Hunteria. 2 (4): 1–9. Проверить значения даты в:
| год =(Помогите) - Норман, Дэвид Б. (1990). «Проблемные тероподы: целурозавры». В Weishampel, Дэвид Б.; Додсон, Питер; Осмольска, Гальска (ред.). Динозаврия. Калифорнийский университет Press. С. 280–305. ISBN 978-0-520-06726-4.
- Ламберт, Дэвид (1993). "Орнитолестес". Лучшая книга динозавров. Дорлинг Киндерсли. стр.78–79. ISBN 978-1-56458-304-8.
- Glut, Дональд Ф. (1997). "Орнитолестес". Динозавры: Энциклопедия. McFarland & Company. С. 643–646. ISBN 978-0-89950-917-4.
- Чуре, Дэниел (1998). "На орбите динозавров теропод" (PDF). Гайя (18): 233–240. Архивировано из оригинал (PDF) на 2011-07-19. Получено 2011-02-06.
- Пол, Грегори С. (2002). «Были ли некоторые динозавры также несветлыми птицами?». Динозавры воздуха: эволюция и потеря полета у динозавров и птиц. Издательство Университета Джона Хопкинса. стр.224–257. ISBN 978-0-8018-6763-7.
- Раухут, Оливер В. (2003). «Оперативные таксономические единицы». Взаимоотношения и эволюция базальных динозавров теропод. Палеонтологическая ассоциация. С. 12–43. ISBN 978-0-901702-79-1.
- Хольц, Томас Р.; Мольнар, Ральф Э.; Карри, Филип Дж. (2004). «Базальная столбняк». В Weishampel, Дэвид Б.; Додсон, Питер; Осмольска, Гальска (ред.). Динозаврия: второе издание. Калифорнийский университет Press. С. 71–110. ISBN 978-0-520-24209-8.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- Фастовский, Давид Э .; Weishampel, Дэвид Б. (2005). "Theropoda I: Природа красная в зубах и когтях". Эволюция и вымирание динозавров. Издательство Кембриджского университета. С. 265–299. ISBN 978-0-521-81172-9.
- Карпентер, Кеннет; Майлз, Клиффорд; Остром, Джон Х.; Клауард, Карен (2005)а). "Переописание малых теропод-манирапторанов Орнитолестес и Целурус"Карпентер, Кеннет (ред.)". Плотоядные динозавры. Жизнь прошлого. Издательство Индианского университета. С. 49–71. ISBN 978-0-253-34539-4. Проверить значения даты в:
| год =(Помогите) - Карпентер, Кеннет; Майлз, Клиффорд; Клауард, Карен (2005)б). "Новый малый теропод из верхнеюрской формации Моррисон штата Вайоминг". В Карпентер, Кеннет (ред.). Плотоядные динозавры. Жизнь прошлого. Издательство Индианского университета. С. 23–48. ISBN 978-0-253-34539-4. Проверить значения даты в:
| год =(Помогите) - Сентер, Фил (2006). "Функция передних конечностей в Ornitholestes hermanni Осборн (Dinosauria, Theropoda) ". Палеонтология. 49 (5): 1029–1034. Дои:10.1111 / j.1475-4983.2006.00585.x.
- Фостер, Джон (2007). «От Гаргантюа до Меньшего: Зверинец Моррисона, Часть II». Юрский Запад: Динозавры формации Моррисон и их мир. Издательство Индианского университета. С. 162–242. ISBN 978-0-253-34870-8.
- Лонг, Джон А.; Схоутен, Питер (2008). "Орнитолесты и родня". Пернатые динозавры: происхождение птиц. Издательство Оксфордского университета. С. 72–77. ISBN 978-0-19-537266-3.
- Пол, Грегори С. (2010). "Ornitholestes hermanni". Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. стр.123–124. ISBN 978-0-691-13720-9.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













