Ячеистая - Alveolate
| Ячеистая | |
|---|---|
 | |
| Ceratium Furca | |
| Научная классификация | |
| Домен: | Эукариоты |
| (без рейтинга): | Потогонные |
| Clade: | ЦАРЬ |
| Clade: | SAR |
| Infrakingdom: | Альвеолаты Кавалер-Смит, 1991 |
| Phyla | |
| Синонимы | |
| |
В альвеолиты (что означает «с полостями»)[2] группа протисты считается основным клады[3] и суперфильм[4] в пределах Эукария, а также называются Альвеолаты.[5]
Характеристики
Наиболее заметной общей характеристикой является наличие корковых (внешних) альвеол (мешочков). Они сплющены пузырьки (мешочки) упакованы сплошным слоем прямо под мембрана и поддерживая его, обычно образуя гибкую пленку (тонкую пленку). В динофлагелляты они часто образуют бронеплиты. Альвеоляты имеют митохондрии с трубчатыми кристами (гребнями), а их жгутики или реснички имеют отчетливое строение.
Практически все секвенированные митохондриальные геномы инфузорий и апикомплексий линейны.[6] Митохондриальный геном Бабезия микроти круглая.[7] Также теперь известно, что этот вид не принадлежит ни к одному из родов. Бабезия или Theileria и для этого нужно будет создать новый род.
Классификация
Альвеолаты включают около 9 основных и второстепенных групп, которые очень разнообразны по форме и, как известно, связаны различными ультраструктурными и генетическими сходствами:[8]
- Инфузории - очень обычные простейшие со многими короткими реснички расположены рядами, и два ядра
- Акавомонидия[8]
- Кольпонемидия[8]
- Динофлагелляты s.l. - в основном морские жгутиконосцы многие из которых имеют хлоропласты
- Perkinsozoa
- Chromerida - морской тип фотосинтетических простейших
- Colpodellida
- Voromonadida
- Apicomplexa - паразитические и вторичные нефотосинтезирующие простейшие, лишенные аксонема конструкции локомотивов, кроме гаметы
Acavomonidia и Colponemidia ранее были сгруппированы как кольпонемиды, а теперь таксон разделен на основе ультраструктурного анализа. Acavomonidia ближе к группе динофлагеллятов / перкинсид, чем Colponemidia.[8] Таким образом, неофициальный термин «кольпонемиды» в его нынешнем виде охватывает две несестринские группы внутри альвеол: Acavomonidia и Colponemidia.[8]
Apicomplexa и динофлагелляты могут быть более тесно связаны друг с другом, чем с инфузориями. Оба имеют пластиды, и у большинства из них имеется пучок или конус микротрубочки в верхней части ячейки. У apicomplexans он образует часть комплекса, используемого для проникновения в клетки-хозяева, тогда как у некоторых бесцветных динофлагеллят он образует стебель, используемый для поедания добычи. К этим двум группам тесно связаны различные другие роды, в основном жгутиконосцы со сходным апикальным строением. К ним относятся свободно живущие члены в Oxyrrhis и Кольпонема, и паразиты в Perkinsus,[9] Parvilucifera, Растримонас и эллобиопсиды. В 2001 году прямое усиление рРНК ген в морском пикопланктон образцы выявили наличие двух новых альвеолятных перепонок, названных группой I и II.[10][11] Группа I не имеет культурных родственников, а группа II связана с паразитом динофлагеллят. Амебофрия, который до сих пор был классифицирован в Syndiniales порядок динофлагеллят.
Связь между некоторыми из этих основных групп была предложена в 1980-х годах, а конкретная взаимосвязь между всеми тремя была подтверждена в начале 1990-х годов генетическими исследованиями, в первую очередь Гаджадхаром. и другие.[12] Кавалер-Смит представила официальное название Alveolata в 1991 году,[5] хотя в то время он фактически считал группировку парафилетический сборка, а не монофилетический группа.
Некоторые исследования показали, что гаплоспориды, в основном паразиты морских беспозвоночных, могут принадлежать сюда, но у них отсутствуют альвеолы, и они теперь находятся среди Cercozoa.
Филогения
На основе сборника следующих работ.[4][13][14]
| Альвеолаты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Недавние исследования показывают, что Dinoflagelllata действительно появилась в Перкинси. Более того, Perkinsea возникла в Apicomplexa.[15]
Таксономия
Альвеолаты Кавалер-Смит 1991 [Альвеолатобионты]
- Тип Цилиофора Дофлейн 1901 стат. п. Коупленд 1956 [Цилиата Перт 1852; Инфузория Bütschli 1887; Реснички, Ciliozoa, Cytoidea, Eozoa, Heterocaryota, Heterokaryota]
- Подтип Постцилиодесматофора Герасимова и Серавин 1976
- Класс Гетеротричея Штейн 1859 г.
- Класс Кариореликтея Корлисс 1974
- Подтип Intramacronucleata Линн 1996
- Класс ?Мезодиния Chen et al. 2015 г.
- Infraphylum Lamellicorticata
- Класс Litostomatea Смолл и Линн 1981
- Класс Armophorea Линн 2004
- Класс Кариакотрихия Орси и др. 2011 г.
- Класс Спиротричея Bütschli 1889 г.
- Infraphylum Ventrata Кавалер-Смит 2004 [Conthreep Линн 2012]
- Порядок ?Discotrichida Chen et al. 2015 г.
- Класс Протокрузия Chen et al. 2015 г. [Protocruziidia de Puytorac, Grain & Mignot 1987]
- Класс Кольподея Смолл и Линн 1981
- Класс Нософорея Смолл и Линн 1981
- Класс Филлофарингея de Puytorac et al. 1974 г.
- Класс Простома Шевяков 1896
- Класс Плагиопилея Смолл и Линн 1985 Сенсю Линн 2008
- Класс Олигогименофорея de Puytorac et al. 1974 г.
- Подтип Постцилиодесматофора Герасимова и Серавин 1976
- Тип Миозоа Кавалер-Смит 1987
- Подтип Colponemidia Тихоненков, Мыльников и Килинг 2013
- Класс Colponemea Кавалер-Смит 1993
- Подтип Acavomonadia Тихоненков и др. 2014 г.
- Класс Acavomonadea Тихоненков и др. 2014 г.
- Подтип Myzozoa Кавалер-Смит 2004
- Инфрафилум Apicomplexa Левин 1970 исправить. Adl et al. 2005 г.
- Порядок ?Витреллида Кавалер-Смит 2017
- Класс ?Myzomonadea Cavalier-Smith & Chao, 2004 г., Sensu Ruggiero et al. 2015 г.
- Класс Chromerea
- порядок Colpodellida Паттерсон и Зельффель, 1991 г. [Spiromonadida Крылов и Мыльников 1986]
- Суперкласс Спорозоа Leuckart 1879 stat. ноя Кавалер-Смит 2013 [Gamontozoa]
- Класс Бластогрегаринида Чаттон и Вильнев, 1936 г. [Бластогрегаринина; Бластогрегаринорина Чаттон и Вильнев, 1936 г.]
- Класс Paragregarea Кавалер-Смит 2014
- Класс Грегариноморфея Грассе 1953 г.
- Класс Кокцидиоморфея Дофлейн 1901
- Инфрафилум Динозоа Кавальер-Смит 1981 исправление. 2003 г.
- Порядок ?Акроцелида Кавалер-Смит и Чао 2004
- Порядок ?Растромонадиды Кавалер-Смит и Чао 2004
- Класс Squirmidea Норен 1999 стат. ноя Кавалер-Смит 2014
- Суперкласс Perkinsozoa Norén et al. 1999 s.s.
- Класс Perkinsea Левин 1978 [Perkinsasida Левин 1978]
- Суперкласс Динофлагеллата Butschli 1885 stat. ноя Кавалер-Смит 1999 Сенсю Кавальер-Смит 2013 [Dinozoa Кавалер-Смит 1981]
- Класс Проноктилуцея
- Класс Эллобиопси Кавалер-Смит 1993 [Ellobiophyceae Лёблих III 1970; Эллобиопсида Whisler 1990]
- Класс Myzodinea Кавалер-Смит 2017
- Класс Оксирея Кавалер-Смит 1987
- Класс Syndinea Чаттон 1920 s.l. [Syndiniophyceae Лёблих III 1970 s.s.; Syndina Кавалер-Смит]
- Класс Endodinea Кавалер-Смит 2017
- Класс Noctiluciphyceae Fensome et al. 1993 г. [Noctilucae Геккель 1866 г.; Noctilucea Haeckel 1866 stat. ноя; Цистофлагеллята Haeckel 1873 stat. ноя Бутшли 1887 г.]
- Класс Dinophyceae Пашер 1914 [Peridinea Эренберг 1830 стат. ноя Wettstein]
- Инфрафилум Apicomplexa Левин 1970 исправить. Adl et al. 2005 г.
- Подтип Colponemidia Тихоненков, Мыльников и Килинг 2013
Развитие
Интересно развитие пластид среди альвеол. Кавалье-Смит предложил альвеолаты, полученные от предка, содержащего хлоропласты, что также привело к появлению Chromista (в хромальвеолят гипотеза). Другие исследователи предположили, что в альвеолах изначально отсутствовали пластиды, и, возможно, динофлагелляты и Apicomplexa приобрели их отдельно. Однако теперь выясняется, что альвеолаты, динофлагелляты, хромрида и гетероконтные водоросли приобрели свои пластиды из красной водоросли, что свидетельствует об общем происхождении этой органеллы во всех этих четырех кладах.[16]
Эволюция
Байесовская оценка помещает эволюцию группы альвеол в ~850 миллион лет назад.[17] Альвеолаты состоят из Myzozoa, Инфузории, и кольпонемиды. Другими словами, термин Myzozoa, означающее «откачивать содержимое добычи», может неформально применяться к общему предку подмножества альвеолят, которые не являются ни инфузориями, ни кольпонемидами. Похищение водорослей является важным фактором эволюции альвеол, так как оно может обеспечить источники эндосимбиоза новых пластид. Период, термин Myzozoa поэтому удобная концепция для отслеживания истории альвеолатного типа.
Предки группы альвеолитов, возможно, были фотосинтетиками.[18] У предковых альвеол, вероятно, был пластида. Chromerids, apicomplexans и перидинин динофлагелляты сохранили это органелла.[19] Если сделать еще один шаг назад, хромериды, перидининовые динофлагелляты и гетероконтные водоросли обладают общим монофилетическим пластидным происхождением, т. е. приобрели свои пластиды от красная водоросль,[16] и поэтому кажется вероятным, что общий предок альвеолят и гетероконтов также был фотосинтезом.
Согласно одной из школ, общий предок динофлагелляты, апикомплекс, Colpodella, Chromerida, и Voromonas был мизоцитотическим хищником с двумя гетеродинамическими жгутики, микропоры, трихоцисты, Rhoptries, микронемы, полярное кольцо и спиральное открытое коноид.[20] Хотя общий предок альвеолят также мог обладать некоторыми из этих характеристик, утверждалось, что мизоцитоз не был одной из этих характеристик, поскольку инфузории поедают добычу по другому механизму.[8]
Продолжающиеся дискуссии касаются количества мембран, окружающих пластиду, у apicomplexans и некоторых динофлагеллят, а также происхождения этих мембран. Этот ультраструктурный признак можно использовать для группировки организмов, и если признак общий, это может означать, что у филов был общий фотосинтетический предок. На основании того, что apicomplexans обладают пластидой, окруженной 4 мембранами, и что перидининовые динофлагелляты обладают пластидой, окруженной 3 мембранами, Petersen et al.[21] были неспособны утверждать, что общая пластида страменопила-альвеол может многократно рециклироваться в типе альвеол, источником которых являются доноры страменопил-альвеолят, через механизм поглощения и эндосимбиоз.
Инфузории представляют собой модельные альвеоляты, которые были глубоко изучены генетически в течение самого длительного периода из всех альвеолистных линий. Они необычны среди эукариот тем, что размножение включает микронуклеус и макронуклеус. Их размножение легко изучается в лаборатории, что исторически сделало их модельными эукариотами. Будучи полностью хищными и лишенными каких-либо остатков пластид, их развитие как филум иллюстрирует, как хищничество и автотрофия[18] находятся в динамическом равновесии и что баланс может колебаться в ту или иную сторону в точке происхождения нового типа от миксотрофных предков, вызывая потерю одной способности.
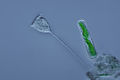
Вортичелла (Цилиофора ) (осталось)

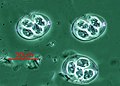
Плазмодий falciparum (Apicomplexa ) в крови




использованная литература
- ^ Li, C.-W .; и другие. (2007). «Ресничные простейшие из докембрийской формации Доушантуо, Венган, Южный Китай». Геологическое общество, Лондон, Специальные публикации. 286: 151–6. Дои:10.1144 / SP286.11.
- ^ «альвеолатный». Словарь / тезаурус Memidex (WordNet). Архивировано из оригинал на 2016-04-11. Получено 2011-01-26.
- ^ Adl, S.M .; и другие. (2012). «Пересмотренная классификация эукариот». Журнал эукариотической микробиологии. 59 (5): 429–514. Дои:10.1111 / j.1550-7408.2012.00644.x. ЧВК 3483872. PMID 23020233.
- ^ а б Руджеро М.А., Гордон Д.П., Оррелл Т.М., Байи Н., Бургуан Т., Бруска Р.С., Кавальер-Смит Т., Гуйри, доктор медицины, Кирк П.М. (2015). «Классификация всех живых организмов более высокого уровня». PLoS ONE. 10 (4): e0119248. Дои:10.1371 / journal.pone.0119248. ЧВК 4418965. PMID 25923521.
- ^ а б Кавальер-Смит, Т. (1991). «Диверсификация клеток у гетеротрофных жгутиконосцев». У Паттерсона, Дэвида Дж .; Ларсен, Джейкоб; Ассоциация систематиков (ред.). Биология свободноживущих гетеротрофных жгутиконосцев. Издательство Оксфордского университета. С. 113–131. ISBN 978-0-19-857747-8.
- ^ Barth, D; Берендонк, Т.У. (2011). "Последовательность митохондриального генома инфузории Paramecium caudatum обнаруживает сдвиг в нуклеотидном составе и использовании кодонов в пределах рода Парамеций". BMC Genomics. 12: 272. Дои:10.1186/1471-2164-12-272. ЧВК 3118789. PMID 21627782.
- ^ Cornillot E, Haddour K, Dassouli A, Noel B, Ranwez V, Vacherie B, Augagneur Y, Brès V, Duclos A, Randazzo S, Carcy B, Debierre-Grockiego F, Delbecq S, Moubri-Ménage K, Shams- Элдин Х, Усмани-Браун С., Бринго Ф, Винкер П., Виварес С. П., Шварц РТ, Шеттерс Т.П., Краузе П.Дж., Горенфлот А., Берри В., Барб В., Бен Мамун С. (2012). "Секвенирование самого маленького апикомплексного генома от человеческого патогена Бабезия микроти". Нуклеиновые кислоты Res. 40 (18): 9102–14. Дои:10.1093 / нар / гкс700. ЧВК 3467087. PMID 22833609.
- ^ а б c d е ж Тихоненков Д.В.; Janouškovec, J; Мыльников АП; Михайлов, К.В. Симдьянов, Т.Г .; Алеошин, В.В.; Килинг, П.Дж. (2014). "Описание Colponema vietnamica sp.n. и Acavomonas peruviana п. ген. п. sp., два новых типа альвеолят (Colponemidia nom. nov. и Acavomonidia nom. nov.) и их вклад в реконструкцию предкового состояния альвеолят и эукариот ». PLoS ONE. 9 (4): e95467. Дои:10.1371 / journal.pone.0095467. ЧВК 3989336. PMID 24740116.
- ^ Чжан, Х; Кэмпбелл, округ Колумбия; Sturm, NR; Дунган, CF; Лин, S (2011). "Сплайсированные лидерные РНК, сдвиги рамки считывания митохондриальных генов и мультибелковая филогения расширяют поддержку этого рода. Perkinsus как уникальная группа альвеолитов ». PLOS ONE. 6 (5): e19933. Дои:10.1371 / journal.pone.0019933. ЧВК 3101222. PMID 21629701.
- ^ Лопес-Гарсия П., Родригес-Валера Ф., Педрос-Алио С., Морейра Д. (2001). «Неожиданное разнообразие мелких эукариот в глубоководном антарктическом планктоне». Природа. 409 (6820): 603–7. Дои:10.1038/35054537. PMID 11214316.
- ^ Мун-ван дер Стаай С.Ю., Де Вахтер Р., Воулот Д. (2001). «Океанические последовательности 18S рДНК из пикопланктона обнаруживают неожиданное эукариотическое разнообразие». Природа. 409 (6820): 607–10. Дои:10.1038/35054541. PMID 11214317.
- ^ Gajadhar, A. A .; и другие. (1991). "Последовательности рибосомной РНК Sarcocystis muris, Theilera annulata, и Crypthecodinium cohnii выявить эволюционные отношения между apicomplexans, динофлагеллятами и инфузориями ». Молекулярная и биохимическая паразитология. 45 (1): 147–153. Дои:10.1016/0166-6851(91)90036-6. PMID 1904987.
- ^ Силар, Филипп (2016), "Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes", HAL Архивы-обзоры: 1–462
- ^ Кавальер-Смит, Томас (5 сентября 2017 г.). «Kingdom Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения». Протоплазма. 255 (1): 297–357. Дои:10.1007 / s00709-017-1147-3. ЧВК 5756292. PMID 28875267.
- ^ Штрассерт, Юрген Ф. Х .; Карнковская, Анна; Хехенбергер, Элизабет; Кампо, Хавьер дель; Колиско, Мартин; Окамото, Норико; Бурки, Фабьен; Яноушковец, Ян; Пуарье, Камилла (2018) [опубликовано в Интернете 10 октября 2017 года]. «Геномика единичных клеток некультивируемых морских альвеол показывает парафилию базальных динофлагеллят». Журнал ISME. 12 (1): 304–308. Дои:10.1038 / ismej.2017.167. ISSN 1751-7370. ЧВК 5739020. PMID 28994824.
- ^ а б Janouskovec, J; Horák, A; Оборник, М; Lukes, J; Килинг, П.Дж. (2010). «Распространенное красное водорослевое происхождение апикомплексана, динофлагеллята и гетероконта пластид». Proc Natl Acad Sci USA. 107 (24): 10949–54. Дои:10.1073 / pnas.1003335107. ЧВК 2890776. PMID 20534454.
- ^ Берни, К; Павловский, Дж (2006). «Молекулярная шкала времени эволюции эукариот, перекалиброванная с учетом непрерывной записи микрофоссилий». Proc Biol Sci. 273 (1596): 1867–72. Дои:10.1098 / rspb.2006.3537. ЧВК 1634798. PMID 16822745.
- ^ а б Рейес-Прието, А; Мустафа, А; Бхаттачарья, Д. (2008). «Множественные гены очевидного водорослевого происхождения предполагают, что инфузории, возможно, когда-то были фотосинтетическими». Curr. Биол. 18 (13): 956–62. Дои:10.1016 / j.cub.2008.05.042. ЧВК 2577054. PMID 18595706.
- ^ Мур Р.Б., Оборник М., Яноусковец Дж., Хрудимски Т., Ванцова М., Грин Д.Х., Райт С.З., Дэвис Н.В., Больх С.Дж., Хейманн К., Слапета Дж., Хуг-Гулдберг О., Логсдон Дж. М., Картер Д.А. (2008). «Фотосинтетические альвеоляты, близкие к апикомплексным паразитам». Природа. 451 (7181): 959–963. Дои:10.1038 / природа06635. PMID 18288187.
- ^ Кувардина, ОН; Леандер, BS; Алешин В.В. Мыльников АП; Килинг, П.Дж.; Симдьянов, Т.Г. (2002). «Филогения кольподеллид (Alveolata) с использованием генных последовательностей малых субъединиц рРНК предполагает, что они являются свободно живущей сестринской группой по отношению к apicomplexans». J Eukaryot Microbiol. 49 (6): 498–504. Дои:10.1111 / j.1550-7408.2002.tb00235.x. PMID 12503687.
- ^ Петерсен Дж, Людвиг А.К., Майкл В., Бунк Б, Ярек М., Борайн Д., Бринкманн Х (2014). "Chromera velia, эндосимбиоз и гипотеза родоплекса - эволюция пластид у криптофитов, альвеолят, страменопилов и гаптофитов (линии CASH) ». Геном Биол Эвол. 6 (3): 666–684. Дои:10.1093 / gbe / evu043. ЧВК 3971594. PMID 24572015.