Agrobacterium tumefaciens - Agrobacterium tumefaciens
| Agrobacterium tumefaciens | |
|---|---|
 | |
| А. tumefaciens прикрепляется к клетке моркови | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Протеобактерии |
| Класс: | Alphaproteobacteria |
| Порядок: | Ризобиальные |
| Семья: | Rhizobiaceae |
| Род: | Ризобий |
| Виды: | Р. radiobacter |
| Биномиальное имя | |
| Rhizobium radiobacter (Бейеринк и ван Делден 1902 г.) Янг и др. 2001 г.[1] | |
| Синонимы | |
| |
Agrobacterium tumefaciens (обновленное научное название Rhizobium radiobacter, синоним Agrobacterium radiobacter)[3][1][4] является причинным агентом коронный галл болезнь (образование опухоли ) более чем у 140 видов эвдикоты. Это стержневидный, Грамотрицательный почва бактерия.[2] Симптомы вызваны введением небольшого сегмента ДНК (известный как Т-ДНК (для «передачи ДНК», не путать с тРНК, которая переносит аминокислоты во время синтеза белка) из плазмиды в клетку растения,[5] который внедряется в завод в полуслучайном месте геном. Геномы растений могут быть созданы с использованием Агробактерии для доставки последовательностей, размещенных в Бинарные векторы Т-ДНК.
А. tumefaciens является альфа-протеобактерии семьи Rhizobiaceae, который включает азотфиксирующий бобовые симбионты. В отличие от азотфиксирующих симбионтов, опухолевидные Агробактерии виды патогенный и не принесут пользы растению. Большое разнообразие растений, пораженных Агробактерии делает его очень важным для сельского хозяйства.[6]
Экономически, А. tumefaciens является серьезным возбудителем грецкие орехи, виноградные лозы, косточковые фрукты, орех деревья сахарная свекла, хрен, и ревень, а стойкий характер опухолей или галлов, вызванных этим заболеванием, делает его особенно вредным для многолетних культур.[7]
А. tumefaciens Оптимально растет при 28 ° C. Время удвоения может варьироваться от 2,5 до 4 часов в зависимости от среды, формата культивирования и уровня аэрации.[8] При температуре выше 30 ° C, А. tumefaciens начинает испытывать тепловой шок, который может привести к ошибкам в делении клеток.[8]
Конъюгация
Быть ядовитый, бактерия содержит опухоль-индуцирующая плазмида (Плазмида Ti или pTi), из 200 kbp, который содержит Т-ДНК и все гены необходимо передать его растительной клетке.[9] Многие штаммы А. tumefaciens не содержат pTi.
Поскольку плазмида Ti важна для того, чтобы вызывать заболевание, события предварительного проникновения в ризосфера придумать для продвижения бактериальная конъюгация - обмен плазмидами между бактериями. В присутствии высказывает мнение, А. tumefaciens производит диффузионный сигнал сопряжения, называемый 30C8HSL или Агробактерии автоиндуктор[нужна цитата ]. Это активирует фактор транскрипции TraR, положительно регулирующий транскрипция генов, необходимых для конъюгации[нужна цитата ].
Способ заражения
А. tumefaciens заражает растение через свою плазмиду Ti. Плазмида Ti интегрирует сегмент своей ДНК, известный как Т-ДНК, в хромосомную ДНК клеток растения-хозяина. А. tumefaciens имеет жгутики которые позволяют ему плавать почва в направлении фотоассимилирует которые накапливаются в ризосфере вокруг корней. Немного напряжения может хемотактически двигаться в сторону химических выделений растений, таких как ацетосирингон и сахара, которые указывают на наличие на растении раны, через которую могут проникнуть бактерии. Фенольные соединения признаны Белок VirA, трансмембранный белок, кодируемый геном virA на плазмиде Ti. Сахара распознаются белком chvE, хромосомным геном-кодируемым белком, расположенным в периплазматическом пространстве.[10]
По крайней мере, 25 генов vir на плазмиде Ti необходимы для индукции опухоли.[нужна цитата ]. Помимо своей роли восприятия, virA и chvE индуцируют другие гены vir. Белок virA имеет автокиназа активность: это фосфорилаты сам по остатку гистидина. Затем белок virA фосфорилирует белок virG по его аспартатному остатку. Белок virG представляет собой цитоплазматический белок, продуцируемый плазмидным геном virG Ti. Это фактор транскрипции, индуцируя транскрипцию вируса опероны. Белок chvE регулирует второй механизм активации генов vir. Повышает чувствительность белка VirA к фенольным соединениям.[10]
Присоединение - это двухэтапный процесс. После первоначального слабого и обратимого прикрепления бактерии синтезируют целлюлоза фибриллы которые прикрепляют их к раненой растительной клетке, к которой они были привлечены. В этом процессе задействованы четыре основных гена: chvA, chvB, pscA, и att. По-видимому, продукты первых трех генов участвуют в собственном синтезе фибрилл целлюлозы. Эти фибриллы также прикрепляют бактерии друг к другу, помогая образовывать микроколония.
VirC, самый важный вирулентный ген, является необходимым шагом в рекомбинации незаконной реколонизации. Он выбирает участок ДНК в растении-хозяине, который будет заменен, и разрезает эту цепь ДНК.
После образования фибрилл целлюлозы кальций-зависимый внешняя мембрана белок вырабатывается рикадгезин, который также способствует прикреплению бактерий к клеточной стенке. Гомологи этого белка можно найти в других ризобиях. В настоящее время существует несколько отчетов о стандартизации протокола опосредованной Agrobacterium трансформации. Влияние различных параметров, таких как время заражения, ацетосирингон, DTT, цистеин, было изучено на сое (Глицин макс)[11]
Возможные растительные соединения, которые инициируют Агробактерии для заражения растительных клеток:[12]
- Ацетосирингон и другие фенольные соединения
- альфа-Гидроксиацетосирингон
- Катехол
- Феруловая кислота
- Галловая кислота
- п-гидроксибензойная кислота
- Протокатеховая кислота
- Пирогалловая кислота
- Резорциловая кислота
- Синапиновая кислота
- Сиринговая кислота
- Ванилин
Формирование Т-образной пилюса
Чтобы перенести Т-ДНК в растительная клетка, А. tumefaciens использует механизм секреции типа IV, включающий производство Т-пилус. При обнаружении ацетосирингона и других веществ преобразование сигнала событие активирует экспрессию 11 генов в VirB оперон которые отвечают за формирование Т-образной пилуса.
Первым образуется пропилин. Это полипептид из 121 аминокислоты, которая требует обработки путем удаления 47 остатков для образования субъединицы Т-пилуса. Субъединица округляется за счет образования пептидная связь между двумя концами полипептида.
Продукты других генов VirB используются для переноса субъединиц через плазматическая мембрана. Дрожжи двугибридные исследования показывают, что VirB6, VirB7, VirB8, VirB9 и VirB10 могут кодировать компоненты транспортера. An АТФаза для активный транспорт подразделений также потребуются.
Перенос Т-ДНК в растительную клетку
B: геном Agrobacterium
C: Ti Плазмида: a: T-ДНК, b: гены Vir, c: происхождение репликации, d: гены катаболизма Opines
D: Растительная клетка
E: митохондрии
F: Хлоропласт
G: ядро
Т-ДНК необходимо вырезать из кольцевой плазмиды. Комплекс VirD1 / D2 разрывает ДНК на левой и правой пограничных последовательностях. Белок VirD2 ковалентно прикреплен к 5'-концу. VirD2 содержит мотив что приводит к тому, что нуклеопротеидный комплекс нацелен на систему секреции типа IV (T4SS).
В цитоплазме клетки-реципиента комплекс Т-ДНК покрывается белками VirE2, которые экспортируются через T4SS независимо от комплекса Т-ДНК.Сигналы ядерной локализации, или NLS, расположенные на VirE2 и VirD2, распознаются белком импортина альфа, который затем связывается с импортином бета и ядерный поровый комплекс перенести Т-ДНК в ядро. VIP1 также, по-видимому, является важным белком в этом процессе, возможно, действующим как адаптер для доставки VirE2 к импортину. Попав внутрь ядра, VIP2 может нацелить Т-ДНК на области хроматин которые активно транскрибируются, так что Т-ДНК может интегрироваться в геном хозяина.
Гены в Т-ДНК
Гормоны
Вызывать желчь образование, Т-ДНК кодирует гены для производства ауксин или индол-3-уксусная кислота через путь IAM. Этот путь биосинтеза не используется во многих растениях для производства ауксина, поэтому это означает, что у растения нет молекулярных средств его регулирования, и ауксин будет вырабатываться конститутивно. Гены производства цитокинины также выражены. Это стимулирует пролиферацию клеток и образование галлов.
Мнения
Т-ДНК содержит гены для кодирования ферменты которые заставляют завод создавать специализированные аминокислота производные, которые бактерии могут метаболизировать, называется высказывает мнение.[13] Мнения представляют собой класс химических веществ, которые служат источником азота для А. tumefaciens, но не для большинства других организмов. Конкретный тип мнения, производимого А. tumefaciens С58 зараженные растения нопалин (Эскобар и другие., 2003).
Две плазмиды Ti нопалинового типа, pTi-SAKURA и pTiC58, были полностью секвенированы. А. tumefaciens C58, первый полностью секвенированный патовар, был впервые выделен из галла кроны вишни. Геном одновременно секвенировал Гуднер. и другие.[14] и дерево и другие.[15] в 2001 году. Геном А. tumefaciens C58 состоит из круглой хромосомы, двух плазмиды, и линейный хромосома. Наличие ковалентно связанной круговой хромосомы является обычным для бактерий, за некоторыми исключениями. Однако наличие как одной кольцевой хромосомы, так и одной линейной хромосомы уникально для группы в этом роде. Две плазмиды - это pTiC58, отвечающая за процессы, участвующие в вирулентность, и pAtC58, получивший название "загадочная" плазмида.[14][15]
Было показано, что плазмида pAtC58 участвует в метаболизме опинов и конъюгируется с другими бактериями в отсутствие плазмиды pTiC58.[16] Если удалить плазмиду pTi, рост опухоли, который является средством классификации этого вида бактерий, не происходит.
Биотехнологическое использование
Возможности передачи ДНК Агробактерии были широко исследованы в биотехнология как средство внедрения чужеродных генов в растения. Марк Ван Монтегю и Джефф Шелл, (Гентский университет и Генетические системы растений, Бельгия ) обнаружил механизм передачи генов между Агробактерии и растений, что привело к разработке методов превращения бактерии в эффективную систему доставки для генная инженерия в растениях.[17] Плазмидная Т-ДНК, которая переносится на растение, является идеальным носителем для генной инженерии.[18] Это делается путем клонирования желаемой генной последовательности в Бинарные векторы Т-ДНК который будет использоваться для доставки интересующей последовательности в эукариотические клетки. Этот процесс был выполнен с использованием firefly люцифераза ген для производства светящихся растений[нужна цитата ]. Эта свечение был полезным прибором при изучении функции хлоропластов растений и в качестве репортерный ген.[19] Также возможно преобразовать Arabidopsis thaliana окунув цветы в бульон Агробактерии: произведенные семена будут трансгенный. В лабораторных условиях Т-ДНК также была перенесена в клетки человека, демонстрируя разнообразие применения вставки.[20]
Механизм, с помощью которого Агробактерии вставляет материалы в клетку-хозяина система секреции типа IV что очень похоже на механизмы, используемые патогены вставлять материалы (обычно белки ) в клетки человека путем секреции типа III. Он также использует тип передачи сигналов, сохраненный у многих грамотрицательных бактерий, который называется проверка кворума[нужна цитата ]. Это делает Агробактерии важная тема медицинских исследований, а также[нужна цитата ].
Естественная генетическая трансформация
Естественная генетическая трансформация в бактерии представляет собой половой процесс, включающий передачу ДНК от одной клетки к другой через промежуточную среду и интеграцию донорной последовательности в геном реципиента посредством гомологичная рекомбинация. А. tumefaciens может подвергаться естественному преобразованию в почве без какой-либо специальной физической или химической обработки.[21]
Цикл болезни
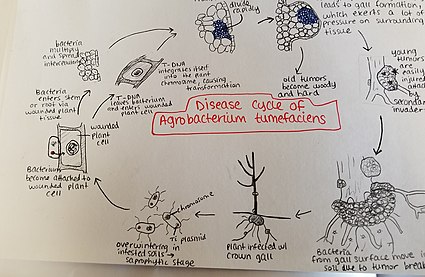
Агробактерии tumefaciens Зимует в зараженных почвах. Агробактерии Виды живут преимущественно сапрофитным образом жизни, поэтому даже паразитирующие на растения виды этого рода обычно выживают в почве в течение длительных периодов времени, даже без присутствия растения-хозяина.[22] Однако при наличии растения-хозяина бактерии проникают в ткань растения через недавние раны или естественные отверстия корней или стеблей у земли. Эти раны могут быть вызваны культивированием, прививкой, насекомыми и т. Д. Как только бактерии попадают в растение, они возникают межклеточно и стимулируют размножение окружающих тканей за счет трансформации клеток. Агробактерии осуществляет этот контроль, вставляя плазмидную Т-ДНК в геном растения. См. Выше для получения более подробной информации о процессе встраивания плазмидной ДНК в геном хозяина. Избыточный рост растительной ткани приводит к образованию галлов на стебле и корнях. Эти опухоли оказывают значительное давление на окружающую растительную ткань, что приводит к раздавливанию и / или деформации этой ткани. Измельчение сосудов приводит к уменьшению потока воды в ксилеме. Молодые опухоли мягкие и поэтому уязвимы для вторичного вторжения насекомых и сапрофитных микроорганизмов. Эта вторичная инвазия вызывает разрушение слоев периферических клеток, а также обесцвечивание опухоли из-за распада. Разрушение мягких тканей приводит к высвобождению Agrobacterium tumefaciens в почву, позволяя возобновить процесс болезни с новым растением-хозяином.[23]
Лечение заболеваний
Болезнь коронной желчи, вызванная Agrobacterium tumefaciens можно контролировать с помощью различных методов. Лучший способ контролировать это заболевание - принимать профилактические меры, такие как стерилизация инструментов для обрезки, чтобы избежать заражения новых растений. Проведение обязательных инспекций питомников и отбраковка зараженных растений, а также отказ от посадки чувствительных растений на зараженных полях также являются ценными методами. Избегать поранения кроны / корней растений во время выращивания важно для предотвращения болезней. В методах садоводства, при которых несколько растений соединяются вместе для роста как одно, например, при бутонизации и прививке.[24] эти методы приводят к травмам растений. Раны являются основным местом попадания бактерий в растение-хозяин. Поэтому рекомендуется выполнять эти техники в течение года, когда Агробактерии не активны. Борьба с корнежевыми насекомыми также помогает снизить уровень инфекции, поскольку эти насекомые вызывают раны (также называемые бактериальными входами) в корнях растений.[23] Рекомендуется сжигать зараженный растительный материал, а не помещать его в компостную кучу, так как бактерии могут жить в почве в течение многих лет.[25]
При лечении этого заболевания также используются методы биологической борьбы. В 1970-х и 1980-х годах обычной практикой обработки проросших семян, сеянцев и подвоев было замачивание их в суспензии K84. K84 состоит из A. radiobacter, который является видом, связанным с А. tumefaciens но не патогенен. K84 продуцирует бактериоцин (агроцин 84), который является антибиотиком, специфичным против родственных бактерий, включая А. tumefaciens. Этот метод, который оказался успешным в борьбе с болезнью в коммерческих масштабах, имел риск передачи K84 своего гена устойчивости патогенным микроорганизмам. Агробактерии. Таким образом, в 1990-х годах был создан генно-инженерный штамм K84, известный как K-1026. Этот штамм так же успешно контролирует коронный галл, как и K84, без необходимости переноса гена устойчивости.[26]
Окружающая среда

Хозяин, окружающая среда и патоген - чрезвычайно важные понятия в отношении патологии растений. Агробактерии иметь самый широкий круг хозяев любого растительного патогена,[27] поэтому главный фактор, который следует учитывать в случае коронкового галла, - это окружающая среда. Существуют различные условия и факторы, которые создают благоприятную среду для А. tumefaciens при заражении различных хозяев. Бактерия не может проникнуть в растение-хозяин без такой точки входа, как рана. Факторы, приводящие к травмам растений, включают культивирование, прививку, травму от замерзания, трещины роста, почвенных насекомых и других животных в окружающей среде, вызывающих повреждение растений. Следовательно, в исключительно суровые зимы часто наблюдается повышенная заболеваемость коронным галлом из-за повреждений, связанных с погодой.[28] Наряду с этим существуют методы опосредования заражения растения-хозяина. Например, нематоды могут действовать как переносчик Агробактерии в корни растений. В частности, паразитирующие на корне нематоды повреждают растительную клетку, создавая рану, через которую проходят бактерии.[29] Наконец, температура является важным фактором при рассмотрении А. tumefaciens инфекция. Оптимальная температура для образования коронного галла из-за этой бактерии составляет 22 градуса по Цельсию из-за термочувствительности переноса Т-ДНК. При более высоких температурах образование опухолей значительно уменьшается.[30]
Смотрите также
использованная литература
- ^ а б Янг JM, Kuykendall LD, Martínez-Romero E, Kerr A, Sawada H (январь 2001 г.). "Пересмотр Rhizobium Frank 1889 с исправленным описанием рода и включением всех видов Agrobacterium Conn 1942 и Allorhizobium undicola de Lajudie et al. 1998 в качестве новых комбинаций: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola и R. vitis ". Международный журнал систематической и эволюционной микробиологии. 51 (Чт 1): 89–103. Дои:10.1099/00207713-51-1-89. PMID 11211278.
- ^ а б Смит EF, Townsend CO (апрель 1907 г.). «Растение-опухоль бактериального происхождения». Наука. 25 (643): 671–3. Bibcode:1907Sci .... 25..671S. Дои:10.1126 / science.25.643.671. PMID 17746161.
- ^ «Rhizobium radiobacter (Agrobacterium tumefaciens) (Agrobacterium radiobacter)». Таксономия UniProt. Архивировано из оригинал на 2011-07-28. Получено 2010-06-30.
- ^ "Браузер таксономии (Agrobacterium radiobacter K84)". Национальный центр биотехнологической информации. Получено 7 декабря 2015.
- ^ Чилтон MD, Драммонд MH, Мерио DJ, Sciaky D, Монтойя А.Л., Гордон MP, Нестер EW (июнь 1977 г.). «Стабильное включение плазмидной ДНК в клетки высших растений: молекулярная основа туморогенеза краун-галла». Ячейка. 11 (2): 263–71. Дои:10.1016/0092-8674(77)90043-5. PMID 890735. S2CID 7533482.
- ^ Мур Л.В., Чилтон В.С., Кэнфилд М.Л. (январь 1997 г.). «Разнообразие опинов и катаболизирующих опин бактерий, выделенных из естественных опухолей коронного желчного пузыря». Прикладная и экологическая микробиология. 63 (1): 201–7. Дои:10.1128 / AEM.63.1.201-207.1997. ЧВК 1389099. PMID 16535484.
- ^ "Коронные галлы". www.missouribotanicalgarden.org. Получено 2019-12-02.
- ^ а б Morton ER, Fuqua C (февраль 2012 г.). «Лабораторное обеспечение агробактерий». Текущие протоколы в микробиологии. Глава 1: Unit3D.1. Дои:10.1002 / 9780471729259.mc03d01s24. ISBN 978-0471729259. ЧВК 3350319. PMID 22307549.
- ^ Гордон Дж. Э., Кристи П. Дж. (Декабрь 2014 г.). «Плазмиды Agrobacterium Ti». Микробиологический спектр. 2 (6). Дои:10.1128 / microbiolspec.PLAS-0010-2013. ЧВК 4292801. PMID 25593788.
- ^ а б Гельвин С.Б. (март 2003 г.). «Трансформация растений, опосредованная Agrobacterium: биология, лежащая в основе инструмента« генной борьбы »». Обзоры микробиологии и молекулярной биологии. 67 (1): 16–37, содержание. Дои:10.1128 / mmbr.67.1.16-37.2003. ЧВК 150518. PMID 12626681.
- ^ Барате П.Л., Кумар Р.Р., Вагмаре С.Г., Павар К.Р., Табе Р.Х. (2018). "Влияние различных параметров на опосредованную Agrobacterium трансформацию в Глицин макс ". Международный журнал перспективных биологических исследований. 8 (1): 99–105.
- ^ Патент США 6483013
- ^ Zupan J, Muth TR, Draper O, Zambryski P (июль 2000 г.). «Перенос ДНК из agrobacterium tumefaciens в растения: праздник фундаментальных открытий». Журнал растений. 23 (1): 11–28. Дои:10.1046 / j.1365-313x.2000.00808.x. PMID 10929098.
- ^ а б Гуднер Б., Хинкль Г., Гаттунг С., Миллер Н., Бланшар М., Куролло Б. и др. (Декабрь 2001 г.). «Последовательность генома растительного патогена и биотехнологического агента Agrobacterium tumefaciens C58». Наука. 294 (5550): 2323–8. Bibcode:2001Sci ... 294.2323G. Дои:10.1126 / science.1066803. PMID 11743194. S2CID 86255214.
- ^ а б Вуд Д. В., Сетубал Дж. К., Каул Р., Монахи Д. Э., Китадзима Дж. П., Окура В. К. и др. (Декабрь 2001 г.). «Геном естественного генного инженера Agrobacterium tumefaciens C58». Наука. 294 (5550): 2317–23. Bibcode:2001Sci ... 294.2317W. CiteSeerX 10.1.1.7.9501. Дои:10.1126 / science.1066804. PMID 11743193. S2CID 2761564.
- ^ Vaudequin-Dransart V, Petit A, Chilton WS, Dessaux Y (1998). "Загадочная плазмида Agrobacterium tumefaciens коинтегрируется с плазмидой Ti и участвует в деградации опина ». Молекулярные взаимодействия растений и микробов. 11 (7): 583–591. Дои:10.1094 / mpmi.1998.11.7.583.
- ^ Шелл Дж, Ван Монтегю М (1977). «Ti-плазмида Agrobacterium tumefaciens, естественный вектор для введения генов nif в растения?». Основные науки о жизни. 9: 159–79. Дои:10.1007/978-1-4684-0880-5_12. ISBN 978-1-4684-0882-9. PMID 336023.
- ^ Zambryski P, Joos H, Genetello C, Leemans J, Montagu MV, Schell J (1983). «Плазмидный вектор Ti для введения ДНК в клетки растений без изменения их нормальной способности к регенерации». Журнал EMBO. 2 (12): 2143–50. Дои:10.1002 / j.1460-2075.1983.tb01715.x. ЧВК 555426. PMID 16453482.
- ^ Корень M (1988). «Светящаяся биотехнология». Бионаука. 38 (11): 745–747. Дои:10.2307/1310781. JSTOR 1310781.
- ^ Куник Т., Цфира Т., Капульник Ю., Гафни Ю., Дингуолл С., Цитовский В. (февраль 2001 г.). «Генетическая трансформация клеток HeLa с помощью Agrobacterium». Труды Национальной академии наук Соединенных Штатов Америки. 98 (4): 1871–6. Bibcode:2001PNAS ... 98.1871K. Дои:10.1073 / pnas.041327598. ЧВК 29349. PMID 11172043.
- ^ Деманеш С., Кей Е., Гурбьер Ф., Симоне П. (июнь 2001 г.). «Естественная трансформация Pseudomonas fluorescens и Agrobacterium tumefaciens в почве». Прикладная и экологическая микробиология. 67 (6): 2617–21. Дои:10.1128 / AEM.67.6.2617-2621.2001. ЧВК 92915. PMID 11375171.
- ^ Schroth MN, Weinhold AR, Mccain AH (март 1971 г.). «Биология и борьба с Agrobacterium tumefaciens». Хильгардия. 40 (15): 537–552. Дои:10.3733 / hilg.v40n15p537.
- ^ а б Агриос Г. Н. (2005). Патология растений (5-е изд.). Амстердам: Elsevier Academic Press. ISBN 9780120445653. OCLC 55488155.
- ^ "Прививочные и окулированные саженцы | Публикации государственного расширения Северной Каролины". content.ces.ncsu.edu. Получено 2017-12-02.
- ^ "Корона галл: Двор и сад: Сад: Расширение Миннесотского университета". www.extension.umn.edu. Архивировано из оригинал на 2017-10-16. Получено 2017-10-15.
- ^ Райдер MH, Джонс DA (1991-10-01). «Биологический контроль коронковой галлы с использованием штаммов Agrobacterium K84 и K1026». Функциональная биология растений. 18 (5): 571–579. Дои:10.1071 / pp9910571.
- ^ "Бактериальный венечный галл фруктовых культур | Огиолин". ohioline.osu.edu. Получено 2017-10-20.
- ^ "Crown Gall - растущее беспокойство на виноградниках". extension.psu.edu. Получено 2017-10-20.
- ^ Карими М., Ван Монтегю М., Гейзен Г. (ноябрь 2000 г.). «Нематоды как векторы для внедрения Agrobacterium в корни растений». Молекулярная патология растений. 1 (6): 383–7. Дои:10.1046 / j.1364-3703.2000.00043.x. PMID 20572986.
- ^ Диллен В., Де Клерек Дж., Капила Дж., Ван Монтегю З.М., Ангенон Дж. (1997-12-01). «Влияние температуры на перенос генов, опосредованный Agrobacterium tumefaciens, растениям». Журнал растений. 12 (6): 1459–1463. Дои:10.1046 / j.1365-313x.1997.12061459.x.
дальнейшее чтение
- Дикинсон М (2003). Молекулярная патология растений. Издательство BIOS Scientific.
- Лай Э.М., Кадо С.И. (август 2000 г.). «Т-пилус Agrobacterium tumefaciens». Тенденции в микробиологии. 8 (8): 361–9. Дои:10.1016 / s0966-842x (00) 01802-3. PMID 10920395.
- Ward DV, Zupan JR, Zambryski PC (январь 2002 г.). «Agrobacterium VirE2 получает лечение VIP1 в ядерном импорте растений». Тенденции в растениеводстве. 7 (1): 1–3. Дои:10.1016 / с1360-1385 (01) 02175-6. PMID 11804814.
- Вебстер Дж, Томсон Дж (1988). «Генетический анализ штамма Agrobacterium Tumefaciens, продуцирующего агроцин, активный против патогена биотипа 3». Молекулярная и общая генетика. 214 (1): 142–147. Дои:10.1007 / BF00340192. S2CID 180063.
внешние ссылки
- Agrobacterium tumefacien Страница генома C58 - секвенировано Cereon Genomics / University of Richmond
| Авторитетный контроль |
|---|