Сульфолобус - Sulfolobus
| Сульфолобус | |
|---|---|
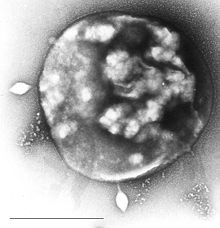 | |
| Электронная микрофотография Сульфолобус заражен Сульфолобус вирус STSV1. Бар = 1 мкм. | |
| Научная классификация | |
| Домен: | |
| Королевство: | |
| Тип: | |
| Класс: | |
| Порядок: | |
| Семья: | |
| Род: | Сульфолобус Брок, Брок, Белли и Вайс, 1972 год. |
| Виды | |
| |
Сульфолобус это род из микроорганизм в семье Sulfolobaceae. Он принадлежит к археи домен.[1]
Сульфолобус виды растут в вулканические источники с оптимальным ростом, происходящим в pH 2-3 и температуры 75-80 ° C, что делает их ацидофилы и термофилы соответственно. Сульфолобус клетки имеют неправильную форму и жгутиковый.
Виды Сульфолобус обычно названы в честь места, из которого они были впервые изолированы, например Sulfolobus solfataricus был впервые изолирован в Сольфатара вулкан. Другие виды можно найти по всему миру в районах вулканической или геотермальной активности, таких как геологические образования, называемые грязевые горшки, которые также известны как Solfatare (множественное число от сольфатары).
Сульфолобус как модель для изучения молекулярных механизмов репликации ДНК
Когда первый геном архей, Метанококк Яннащий, была полностью секвенирована в 1996 г., было обнаружено, что гены в геноме Methanococcus jannaschii участвующие в репликации, транскрипции и трансляции ДНК были больше связаны со своими аналогами у эукариот, чем с таковыми у других прокариот. В 2001 году первая последовательность генома Сульфолобус, Sulfolobus solfataricus P2, был опубликован. В геноме P2 гены, связанные с репликацией хромосом, также оказались более близкими к генам эукариот. Эти гены включают ДНК-полимеразу, примазу (включая две субъединицы), MCM, CDC6 / ORC1, RPA, RPC и PCNA. В 2004 году происхождение репликации ДНК Sulfolobus solfataricus и Sulfolobus acidocaldarius были идентифицированы. Он показал, что оба вида содержат в своем геноме два источника. Это был первый случай, когда в прокариотической клетке использовалось более одного ориджина репликации ДНК. Механизм репликации ДНК у архей эволюционно консервативен и подобен таковому у эукариот. Сульфолобус сейчас используется в качестве модели для изучения молекулярных механизмов репликации ДНК у архей. А поскольку система репликации ДНК у архей намного проще, чем у эукариот, было высказано предположение, что археи можно использовать в качестве модели для изучения гораздо более сложной репликации ДНК у эукариот.
Роль в биотехнологии
Сульфолобус белки представляют интерес для биотехнологии и промышленного использования из-за их термостабильный природа. Одно из приложений - создание искусственных производных от S. acidocaldarius белки, названные аффитины. Однако внутриклеточные белки не обязательно стабильны при низком pH, поскольку Сульфолобус виды поддерживают значительный градиент pH через внешнюю мембрану. Sulfolobales метаболически зависят от серы: гетеротрофный или автотрофный, их энергия исходит от окисление из сера и / или клеточное дыхание в котором сера действует как конечный акцептор электронов. Например, S. tokodaii известно окислять сероводород к сульфат внутриклеточно.
Статус генома
Полный геномы были упорядочены для S. acidocaldarius DSM 639 (2,225,959 нуклеотиды ),[2] S. solfataricus P2 (2 992 245 нуклеотидов),[3] и S. tokodaii ул. 7 (2 694 756 нуклеотидов).[4]
Структура генома
Археон Sulfolobus solfataricus имеет круговую хромосому, состоящую из 2 992 245 п.н. Другой упорядоченный вид, S. tokodaii также имеет круговую хромосому, но немного меньше - 2 694 756 п.н. У обоих видов отсутствуют гены ftsZ и minD, что характерно для секвенированных Crenarchaeota. Они также кодируют цитрат-синтазу и две субъединицы 2-оксикислоты: ферредоксин оксидоредуктазу, которая играет ту же роль, что и альфа-кетоглутаратдегидрогеназа в цикле TCA (трикарбоновая кислота / Кребса / лимонная кислота). Это указывает на то, что Сульфолобус имеет систему цикла TCA, аналогичную той, что обнаружена в митохондриях эукариот. Другие гены в дыхательной цепи, которые участвуют в производстве АТФ, не были похожи на гены, обнаруженные у эукариот. Цитохром с - один из таких примеров, который играет важную роль в переносе электронов к кислороду у эукариот. Это также было обнаружено у A. pernix K1. Поскольку этот шаг важен для таких аэробных микроорганизмов, как Сульфолобус, вероятно, он использует другую молекулу для той же функции или имеет другой путь.
Структура клетки и метаболизм
Сульфолобус может расти литоавтотрофно за счет окисления серы или хемогетеротрофно с использованием серы для окисления простых восстановленных углеродных соединений. Однако гетеротрофный рост наблюдается только в присутствии кислорода. Основные метаболические пути - это гликолитический путь, пентозофосфатный путь и цикл TCA.
У всех архей есть липиды с эфирными связями между головной группой и боковыми цепями, что делает липиды более устойчивыми к нагреванию и кислоте, чем липиды, связанные сложным эфиром бактерий и эукариот. Sulfolobales известны необычными тетраэфирными липидами. У Sulfolobales липиды, связанные с эфиром, ковалентно связаны через «бислой», образуя тетраэфиры. Таким образом, технически тетраэфиры образуют монослой, а не бислой. Тетраэфиры помогают Сульфолобус виды выживают как в экстремальной кислоте, так и в высокой температуре.
Экология
S. solfataricus был обнаружен в разных областях, включая Йеллоустонский Национальный Парк, Mount St. Helens, Исландия, Италия, и Россия назвать несколько. Сульфолобус находится почти везде, где есть вулканическая активность. Они хорошо себя чувствуют в среде с температурой около 80 ° C, pH около 3 и присутствием серы. Другой вид, S. tokodaii, был расположен в кислотном спа в Горячие источники Беппу, Кюсю, Япония. В отложениях от ~ 90 м ниже морского дна на окраине перуанского материка преобладают интактные архейские тетраэфиры, а значительная часть сообщества - это осадочные археи, таксономически связанные с кренархеальными Sulfolobales (Sturt, и другие., 2004).
Ответ на повреждение ДНК
Контакт с Sulfolobus solfataricus или Sulfolobus acidocaldarius к агентам, повреждающим ДНК. УФ-облучение, блеомицин или митомицин С вызывают агрегацию клеток.[5][6] Другие факторы физического стресса, такие как изменение pH или температуры, не вызывают агрегации, что позволяет предположить, что индукция агрегации вызвана именно повреждением ДНК.[6] Ajon et al.[5] показали, что УФ-индуцированная агрегация клеток опосредует обмен хромосомными маркерами с высокой частотой в S. acidocaldarius. Скорости рекомбинации превышали таковые в неиндуцированных культурах до трех порядков. Wood et al.[7] также показали, что УФ-облучение увеличивает частоту рекомбинации из-за генетического обмена в S. acidocaldarius. Frols et al.[6][8] и Ajon et al.[5] предположили, что процесс переноса ДНК, индуцируемый ультрафиолетом, и последующая гомологичная рекомбинационная репарация представляют собой важный механизм для поддержания целостности хромосом в S. acidocaldarius и S. solfataricus. Этот ответ может быть примитивной формой полового взаимодействия, подобной более хорошо изученной бактериальной трансформации, которая также связана с переносом ДНК между клетками, приводящим к гомологичной рекомбинационной репарации повреждений ДНК.[9][10]
В UPS оперон
В UPS оперон из Сульфолобус вид сильно индуцируется УФ-облучение. В пили кодируемые этим опероном, используются для стимулирования клеточной агрегации, которая необходима для последующего обмена ДНК между клетками, что приводит к гомологичная рекомбинация. Исследование Sulfolobales acidocaldarius UPS оперон показал, что один из генов оперона, saci-1497, кодирует эндонуклеаза III, который разрезает поврежденную УФ излучением ДНК; и еще один ген оперона, saci-1500, кодирует RecQ-подобная геликаза который способен расслабиться гомологичная рекомбинация промежуточные звенья, такие как Праздничные развязки.[11] Было высказано предположение, что Saci-1497 и Saci-1500 действуют в рамках механизма репарации ДНК на основе гомологичной рекомбинации, который использует перенесенную ДНК в качестве матрицы.[11] Таким образом, считается, что UPS система в сочетании с гомологичная рекомбинация обеспечивают реакцию на повреждение ДНК, которая спасает Sulfolobales от угроз, повреждающих ДНК.[11]
Сульфолобус как вирусный хозяин
Лизогенный вирусы заражают Сульфолобус для защиты. Вирусы не могут выжить в чрезвычайно кислых и жарких условиях, которые Сульфолобус живет в, и поэтому вирусы используют Сульфолобус как защита от суровых погодных условий. Эта связь позволяет вирусу размножаться внутри архей, не разрушаясь окружающей средой. Вирусы Sulfolobus относятся к умеренным или постоянным лизогенам. Постоянные лизогены отличаются от лизогенных бактериофагов тем, что клетки-хозяева не лизируются после индукции продукции Fuselloviridae и в конечном итоге возвращаются в лизогенное состояние. Они также уникальны в том смысле, что гены, кодирующие структурные белки вируса, постоянно транскрибируются и, по-видимому, индуцируется репликация ДНК. Вирусы, заражающие архей, такие как Sulfolobus, должны использовать стратегию, чтобы избежать длительного прямого воздействия того типа среды, в которой живет их хозяин, что может объяснить некоторые из их уникальных свойств.
Смотрите также
использованная литература
- ^ Увидеть NCBI веб-страница на Sulfolobus. Данные извлечены из "Ресурсы таксономии NCBI". Национальный центр биотехнологической информации. Получено 2007-03-19.
- ^ Чен, L; Брюггер, К; Сковгаард, М; Реддер, П; Она, Q; Torarinsson, E; Греве, B; Awayez, M; Зибат, А; Кленк, HP; Гарретт, РА (июль 2005 г.). «Геном Sulfolobus acidocaldarius, модельного организма Crenarchaeota». Журнал бактериологии. 187 (14): 4992–9. Дои:10.1128 / JB.187.14.4992-4999.2005. ЧВК 1169522. PMID 15995215.
- ^ Она, Q; Сингх, РК; Confalonieri, F; Zivanovic, Y; Allard, G; Awayez, MJ; Chan-Weiher, CC; Clausen, IG; Curtis, BA; Де Моорс, А; Erauso, G; Флетчер, К; Гордон, премьер-министр; Хейкамп-де Йонг, я; Джеффрис, AC; Козера, CJ; Медина, N; Пэн, X; Ти-Нгок, HP; Реддер, П; Schenk, ME; Терио, С; Толструп, Н; Шарлебуа, Р.Л .; Дулиттл, ВФ; Дюге, М; Gaasterland, T; Гарретт, РА; Раган, Массачусетс; Sensen, CW; Ван дер Ост, Дж (3 июля 2001 г.). «Полный геном кренархея Sulfolobus solfataricus P2». Труды Национальной академии наук Соединенных Штатов Америки. 98 (14): 7835–40. Дои:10.1073 / pnas.141222098. ЧВК 35428. PMID 11427726.
- ^ Каварабаяси, Й; Хино, Y; Хорикава, Н; Джин-но, К; Такахаши, М; Секин, М; Баба, S; Анкай, А; Kosugi, H; Хосояма, А; Фукуи, S; Нагаи, Y; Нисидзима, К; Otsuka, R; Накадзава, Н; Такамия, М; Като, Y; Йошизава, Т; Танака, Т; Kudoh, Y; Ямазаки, Дж; Kushida, N; Огучи, А; Аоки, К; Масуда, S; Янагий, М; Нисимура, М; Ямагиши, А; Осима, Т; Кикучи, H (31 августа 2001 г.). «Полная последовательность генома аэробного термоацидофильного кренархея, штамм Sulfolobus tokodaii7». ДНК исследования. 8 (4): 123–40. Дои:10.1093 / днарес / 8.4.123. PMID 11572479.
- ^ а б c Ajon M; Fröls S; van Wolferen M; и другие. (Ноябрь 2011 г.). «УФ-индуцируемый обмен ДНК в гипертермофильных архее, опосредованный пилями типа IV» (PDF). Мол. Микробиол. 82 (4): 807–17. Дои:10.1111 / j.1365-2958.2011.07861.x. PMID 21999488.
- ^ а б c Fröls S; Ajon M; Вагнер М; и другие. (Ноябрь 2008 г.). «Индуцируемая УФ-излучением клеточная агрегация гипертермофильной археи Sulfolobus solfataricus опосредована образованием пилей» (PDF). Мол. Микробиол. 70 (4): 938–52. Дои:10.1111 / j.1365-2958.2008.06459.x. PMID 18990182.
- ^ Вуд ER; Ghané F; Grogan DW (сентябрь 1997 г.). «Генетические реакции термофильных архей Sulfolobus acidocaldarius на коротковолновый УФ-свет». J. Bacteriol. 179 (18): 5693–8. Дои:10.1128 / jb.179.18.5693-5698.1997. ЧВК 179455. PMID 9294423.
- ^ Fröls S; Белый MF; Шлепер C (февраль 2009 г.). «Реакции на УФ-повреждение у модельного архея Sulfolobus solfataricus». Biochem. Soc. Транс. 37 (Pt 1): 36–41. Дои:10.1042 / BST0370036. PMID 19143598.
- ^ Брутто J; Бхаттачарья Д. (2010). «Объединение пола и происхождения эукариот в развивающемся кислородном мире». Биол. непосредственный. 5: 53. Дои:10.1186/1745-6150-5-53. ЧВК 2933680. PMID 20731852.
- ^ Bernstein, H; Бернштейн, С (2010). «Эволюционное происхождение рекомбинации во время мейоза». Бионаука. 60 (7): 498–505. Дои:10.1525 / bio.2010.60.7.5.
- ^ а б c ван Вольферен М, Ма Х, Альберс С.В. (2015). «Белки процессинга ДНК, участвующие в УФ-индуцированной стрессовой реакции сульфолобалов». J. Bacteriol. 197 (18): 2941–51. Дои:10.1128 / JB.00344-15. ЧВК 4542170. PMID 26148716.
- Мэдиган М; Мартинко Дж, ред. (2005). Биология микроорганизмов Брока (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7.
дальнейшее чтение
Научные журналы
- Судебная комиссия Международного комитета по систематике прокариот (2005 г.). "Номенклатурными типами отрядов Acholeplasmatales, Halanaerobiales, Halobacteriales, Methanobacteriales, Methanococcales, Methanomicrobiales, Planctomycetales, Prochlorales, Sulfolobales, Thermococcales, Thermoproteales и Verrucomicrobiaces, являются роды Methanobachanobacteria, Methanobachanobacteria, Methanobachanicrobiales, Methanobacherobales и Verrucomicrobiales, Methanobachanterobiales, Methanobachanterobiales, Methanobachanterobaccus, Methanobachanterobiales, Methanobachanterobaccusus, Methanochanoplacterobiales, Methanobachanoplasterobiales, Methanobachanoplacterobiales, Methanochanoplacterobiales, Metharaobachanoplacterobiales, Methanochanoplacterobacusus, Methylum, Methanochanoplacterobaccus, Methara ... , Thermococcus, Thermoproteus и Verrucomicrobium, соответственно. Мнение 79 ". Int. J. Syst. Evol. Микробиол. 55 (Pt 1): 517–518. Дои:10.1099 / ijs.0.63548-0. PMID 15653928.
- Брок ТД; Брок К.М.; Живот RT; Вайс Р.Л. (1972). «Sulfolobus: новый род сероокисляющих бактерий, живущих при низком pH и высокой температуре». Arch. Микробиол. 84 (1): 54–68. Дои:10.1007 / BF00408082. PMID 4559703.
Научные книги
- Стеттер, К.О. (1989). «Порядок III. Sulfolobales ord. Nov. Семейство Sulfolobaceae fam. Nov.». В JT Staley; Депутат Брайант; Н. Пфенниг; JG Holt (ред.). Руководство Берджи по систематической бактериологии. 3 (1-е изд.). Балтимор: Williams & Wilkins Co., стр. 169.
Научные базы данных
внешние ссылки
- Страница таксономии NCBI для Sulfolobus
- Поиск на страницах таксономии "Древо жизни" для Sulfolobus
- Страница поиска видов2000 для Sulfolobus
- Страница MicrobeWiki для Sulfolobus
- Страница LPSN для Sulfolobus
- Сравнительный анализ геномов Sulfolobus (в DOE Система IMG )
- Проекты Sulfolobus Genome (из База данных Genomes OnLine )