Эпителиоидная клетка - Epithelioid cell
Эта статья нужны дополнительные цитаты для проверка. (Август 2015 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |

Согласно общепринятой точке зрения эпителиоидные клетки (также называемый эпителиоидным гистиоциты ) являются производными от активированных макрофаги напоминающий эпителиальные клетки.[1][2][3]
Структура и функции

Структурно эпителиоидные клетки (при исследовании с помощью световой микроскопии после окрашивания гематоксилином и эозином) имеют удлиненную форму с мелкозернистой бледной эозинофильной (розовой) цитоплазмой и центральными яйцевидными ядрами (овальными или удлиненными), которые менее плотны, чем у а лимфоцит.[4] Они имеют нечеткую форму и часто сливаются друг с другом, образуя агрегаты, известные как гигантские клетки. При рассмотрении просвечивающая электронная микроскопия в эпителиоидных клетках в области пластинчатого комплекса Гольджи выявляются не только зонированные, но и гладкие пузырьки с плотным центром, а также большое количество (более 100) крупных гранул диаметром до 340 нм и с мелкозернистым матриксом более светлым, чем в макрофагах. гранулы, иногда с перигранулярным ореолом. «Самая заметная особенность этих камер - огромная площадь Гольджи; может присутствовать до 6 отдельных стопок цистерн Гольджи, а также несколько покрытых щетиной и многочисленных гладких везикул ».[5][4] Эпителиоидные клетки имеют плотно пересекающиеся клеточные мембраны в виде «застежек-молний», которые соединяют соседние клетки.[3] Эти клетки играют центральную роль в формировании гранулем, которые связаны со многими серьезными заболеваниями.[4] В гранулемах эпителиоидные клетки выполняют функции разграничения.[3]
Особенности цитоскелета

Показано, что эпителиоидная клетка цитоскелет образованный филаментами, существенно отличается от цитоскелета макрофагов.[6][7] В этих клетках происходит значительное увеличение количества филаментов, где филаменты (от 90 до 100 A) окружают цитоцентр в виде характерного кольцевого пучка, часто ответвляющегося в цитоплазму. Благодаря такому цитоскелету смежные эпителиоидные клетки обнаруживают сложную цитоплазматическую перекрестную перемычку.[6] С помощью методов быстрого = замораживания и замораживания-замещения (быстрое замораживание, проникающее травление и замораживание-замещение) было показано, что организации трехмерной метаструктуры цитоскелета эпителиоидных клеток, образующихся в фокусе гранулематозный воспаление, более совместимый с цитоскелетом, характерным для типичной эпителиальной клетки, чем с цитоскелетом активных и подвижных макрофагов. Показано, что плотные сети промежуточных волокон, связанных с ядрами, митохондриями и другими органеллами, контролируются повсюду в цитоплазме эпителиоидных клеток. Некоторые пучки актинических филаментов располагались в филоподиях ниже мембран клеток. Было ясно продемонстрировано точное межпальцевое утроение мембран клеток между взаимодействующими эпителиоидными клетками. Выявлены межпальцевые филоподии. Характерным признаком эпителиоидных клеток является их агрегация с образованием плотных межпальцевых тройников в виде застежки «молния», которая, по-видимому, может иметь важное значение для образования плотной зоны отграничения тела от возбудителя при образовании эпителиоидной клетки. гранулемы.[7]
Иммунологический фенотип и иммунологические маркеры
При использовании антител к антигенам RFD9, RFD7 и HLA-DR было обнаружено, что все эпителиоидные клетки имеют иммунологический фенотип RFD9 + / RFD7- / HLA-DR +.[8] Серия моноклональные антитела Были получены IHY-1, IHY-2, IHY-3, которые можно использовать для точной идентификации эпителиальных клеток, образующихся при этиологически различных формах гранулематозного воспаления. Антитело IHY-1 реагирует с эпителиоидными клетками в саркоид гранулемы, а также эпителиоидные клетки различных гранулематозных заболеваний, включая туберкулез. IHY-2 и IHY-3 l антитела, моноклональные антитела, реагируют с эпителиоидными клетками в саркоидоз но не в туберкулез.[9]
Клиническое значение
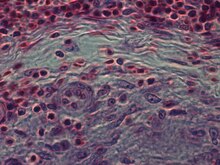
Эпителиоидные клетки являются важной характеристикой эпителиоидных клеток. гранулемы.[10] Эпителиоидно-клеточная гранулема может быть определена как специфически и структурно организованная совокупность эпителиоидных клеток, макрофагов, лимфоцитов и дендритных клеток. Гранулемы инородного тела можно рассматривать как организованный набор макрофагов, включающий простые скопления гигантские клетки окружающие инертные вещества, такие как шовный материал, - так называемые «неиммунные гранулемы». Образование гранулем связано с патогенами, которые научились уклоняться от иммунной системы хозяина различными способами, такими как сопротивление фагоцитоз и убийство внутри макрофагов. Неусвояемость вещества макрофагами - частый признак гранулематозного воспаления.[4] Гранулемы пытаются отгородить эти организмы и предотвратить их дальнейший рост и распространение. Исторически распространенные и деструктивные заболевания, такие как туберкулез, проказа и сифилис гранулематозные состояния. Формирование гранулемы также является признаком многих современных заболеваний, таких как грибковые инфекции, саркоидоз и другие заболевания. болезнь Крона.[4]
История научных исследований
Первое упоминание об эпителиоидных клетках как об особой клеточной форме произошло в 19 веке в работах А. Koch R и Корнил Дж., который верил лейкоциты быть зачинателями эпителиоидных клеток туберкулеза. В опытах на кроликах Ерсин А (1988) и Боррель А (1893) показали, что эпителиоидные клетки образуются из мононуклеарных лейкоцитов крови.[11] Основные закономерности образования эпителиоидных клеток были впервые описаны в первой половине 20 века Lewis M (1925). Этот исследователь показал, что кровь моноциты в клеточные культуры лейкоцитов смешанной крови птиц (взятых от взрослых птиц, а также от эмбрионов разного возраста), мышей и людей при культивировании in vitro, трансформируются в типичные макрофаги и эпителиоидные клетки с последующим образованием гигантских многоядерных клеток. Образование клеток эпителиоидного типа было отмечено Lewis M на 2-3-й день культивирования лейкоцитов.[12] Позднее в исследовании аналогичного плана Jerry S и Weiss L (1966) при использовании культур смешанных лейкоцитов крови курицы (отделенных от сердечной крови Rhode Island Red) и электронной микроскопии показали, что трансформация моноцитов курицы в эпителиоидные клетки начинают культивироваться через 3–4 дня и заканчиваются через 5–6 дней.[6] Поскольку все предыдущие исследователи указывали, что эпителиоидные клетки образуются из моноцитов, а моноциты и макрофаги были объединены в единый система мононуклеарных фагоцитов, Ван Фурт и др. (1972), ссылаясь на работу Sutton J и Weiss L (1966),[6] формально относили эпителиоидные клетки к системе мононуклеарных фагоцитов.[2] Однако они не указали точно, из каких клеток системы мононуклеарных фагоцитов происходят эпителиоидные клетки. В то же время они сделали очень осторожный вывод, что «эпителиоидные клетки, встречающиеся в этих поражениях, также возникают из моноцитов или макрофагов».[2] Адамс Д. (1976), полагая, что эпителиоидные клетки являются заключительной стадией клеточной дифференцировки клеток мононуклеарной системы фагоцитов, сформулировал концепцию цитоморфогезиса эпителиоидных клеток, согласно которой эпителиоидные клетки рассматриваются как производные активированных макрофагов (которые большинство исследователей по-прежнему придерживаются этого мнения). Не основан на предположении, что «стимуляция макрофагов созревает дальше в незрелые эпителиоидные клетки и в конечном итоге в зрелых».[13] Ри и др. (1979) в экспериментах на крысах с помощью метода электронной микроскопии показали, что одной из основных цитоморфологических особенностей эпителиоидных клеток, отличающих эти клетки от макрофагов, является наличие в них характерных специфических гранул, которые они назвали гранулами эпителиоидных клеток. На основании собственных данных они поддержали концепцию цитоморфогенеза эпителиоидных клеток, согласно которой эпителиоидные клетки рассматриваются как производные активированных макрофагов.[5]Позже Turk JL и Narayanan RB (1982) предложили в исследовании различать два типа эпителиоидных клеток: «везикулярный» и "секреторный" эпителиоидные клетки. Предполагается, что «везикулярные эпителиоидные клетки могут развиваться из« секреторных »эпителиоидных клеток в процессе дегенерации.[10] В поисках иммунологических механизмов, влияющих на формирование эпителиальных клеток, Cipriano et al. (2003) получили данные, указывающие на возможное влияние Ил-4 о формировании фенотипа макрофагов, сходного с фенотипом эпителиоидных клеток.[14] Однако не все результаты исследований, посвященных изучению закономерностей и механизмов цитоморфогенеза эпителиоидных клеток, укладываются в представления о происхождении эпителиоидных клеток из макрофагов. Deimann J и Fahimi H (1980) показали, что эпителиоидные клетки в гранулемах, индуцированных в печени крыс инъекцией глюкана, бета-1,30-полиглюкозы, образуются не из Клетки Купфера - зрелый дифференцированный макрофаги, но из моноцитов крови.[15]De Vos et al. (1990) получили данные, позволяющие предположить, что в очагах гранулематозного воспаления, в гранулематозном лимфаденит, эпителиоидные клетки образуются не из дифференцированных макрофагов, а из так называемых плазмацитоид моноциты (имеют сходство с плазмацитами). Это дополнительно подтверждается ультраструктурным сходством между плазматическими моноцитами и эпителиоидными клетками. Настоящее ультраструктурное и иммуноэлектронно-микроскопическое исследование гранулем эпителиоидных клеток предоставило дополнительные аргументы в пользу этой гипотезы.[16] Архипов С (1997, 2012)[17][18] с использованием культур клеток брюшины, лейкоцитов крови и кости костный мозг клеток мышей, показали, что макрофаги и эпителиоидные клетки образуются из разных типов моноцитов. Было показано, что эпителиоидные клетки образуются только из моноцитов плазмоцитоидного типа, названного преэпителиоидными клетками, минуя стадию дифференциация в макрофаги. Было показано, что при хроническом воспалении количество преэпителиоидных моноцитарных клеток, коммитируемых при дифференцировке эпителиоидных клеток, увеличивается в очаге воспаления, в крови и костном мозге. используя мышь Инбредные линии, в отличие от восприимчивости к Микобактерии туберкулеза, было показано, что количество преэпителиоидных моноцитарных клеток, образующихся при хроническом воспалении, определяется генетически. Полученные результаты показали, что морфогенез эпителиоидно-клеточных гранулем может определяться разными стартовыми генетически детерминированный уровень пула преэпителиоидных клеток моноцитоидного типа, их перетекание в очаг гранулематозного воспаления, интенсивность их дифференцировки в эпителиоидные клетки, минуя стадию дифференцировки в макрофаги, и их эндомитоз Мероприятия.[18]
Смотрите также
Рекомендации
- ^ «Оксфордский справочник: эпителиоидная клетка».
- ^ а б c Ван Фурт Р., Кон З. А., Хирш Дж. Г., Хамфри Дж. Х., Спектор В. Г., Лангевурт Х. Л. (1972). «Система мононуклеарных фагоцитов: новая классификация макрофагов, моноцитов и их клеток-предшественников». Орган здоровья Bull World. 46 (6): 845–52. PMID 4538544.
- ^ а б c Рамакришнан Л. (апрель 2012 г.). «Возвращаясь к роли гранулемы при туберкулезе». Нат Рев Иммунол. 12 (5): 352–66. Дои:10.1038 / nri3211. PMID 22517424.
- ^ а б c d е Cross S (30.04.2018). Патология Андервуда: клинический подход. Кросс, Саймон С. (Седьмое изд.). Эдинбург. ISBN 9780702072123. OCLC 1040695915.
- ^ а б Ван дер Ри HJ, Ван дер Бург-де Винтер CP, Daems WT (апрель 1979 г.). «Дифференциация моноцитов в макрофаги, эпителиоидные клетки и многоядерные гигантские клетки в подкожных гранулемах II. Пероксидатическая активность». Исследования клеток и тканей. 197: 355–78. Дои:10.1007 / BF00233564.
- ^ а б c d Саттон Дж. С., Вайс Л. (1966). «Превращение моноцитов в тканевой культуре в макрофаги, эпителиоидные клетки и многоядерные гигантские клетки. Исследование под электронным микроскопом». J Cell Biol. 28 (2): 303–32. Дои:10.1083 / jcb.28.2.303. PMID 5914695.
- ^ а б Баба Т., Сакагути Н., Хотчи М., Оно С. (1992). «Трехмерное исследование эпителиоидных клеток методом быстрого замораживания и глубокого травления в гранулемах, индуцированных мурамилдипептидом». Вирхов Архив B. 63: 63–70. Дои:10.1007 / bf02899245. PMID 1362023.
- ^ Манро С.С., Кэмпбелл Д.А., Коллингс Л.А., Поултер Л.В. (1987). «Моноклональные антитела различают макрофаги и эпителиоидные клетки при саркоидозе и лепре». Клин Эксп Иммунол. 68 (2): 282–87. PMID 3308212.
- ^ Ишиока С., Вивиен Х.В., Хияма К., Маэда А., Ямакидо М. (1999). «Новые моноклональные антитела против эпителиоидных клеток саркоидных гранулем». Exp Lung Res. 25 (8): 663–70. Дои:10.1080/019021499269981. PMID 10643563.
- ^ а б Turk JL, Narayanan RB (апрель 1982 г.). «Происхождение, морфология и функция эпителиоидных клеток». Иммунобиология. 161 (3–4): 274–82. Дои:10.1016 / S0171-2985 (82) 80083-1. PMID 7047374.
- ^ Максимов А. "Роль некорнулярных лейкоцитов крови в формировании бугорка", журнал Infectious Disease, 1925, 37 (5): 418–29. [1]
- ^ Льюис MR (1925). «Формирование макрофагов, эпителиоидных клеток и гигантских клеток из лейкоцитов в инкубированной крови». Am J Pathol. 1 (1 Am J Pathol): 91–100. PMID 19969635.
- ^ Адамс Д.О. (1976). «Гранулематозный воспалительный ответ. Обзор». Am J Pathol. 84 (1): 164–91. PMID 937513.
- ^ Чиприано И.М., Мариано М., Фреймуллер Э, Уитакер-Карнейро С. Р. (2003). «Мышиные макрофаги, культивируемые с IL-4, приобретают фенотип, подобный фенотипу эпителиоидных клеток от гранулематозного воспаления». Воспаление. 27 (4): 201–11. Дои:10.1023 / а: 1025084413767. PMID 14527173.
- ^ Дейманн В., Фахими HD (1980). «Гранулемы печени, индуцированные глюканом. Ультраструктурное и пероксидазно-цитохимическое исследование». Lab Invest. 43 (2): 172–81. PMID 7401631.
- ^ Де Вос Р., Де Вольф-Петерс С., Факкетти Ф, Десмет В. (1990). «Плазмацитоидные моноциты в эпителиоидно-клеточных гранулемах: ультраструктурное и иммуноэлектронное микроскопическое исследование». Ультраструктура Патол. 14 (4): 291–302. Дои:10.3109/01913129009032244. PMID 2382309.
- ^ Архипов, С (1997). Эпителиоидная клетка: новая концепция происхождения и дифференциации (PDF). НОВОСИБИРСК: "НАУКА" СИБИРСКОЕ ПРЕДПРИЯТИЕ РАН. С. 1–88. ISBN 5-02-031381-5.
- ^ а б Архипов, С (2012). «Эпителиоидная клетка: новое мнение о ее природе, происхождении, гистогенезе, цитоморфогенезе, морфофункциональной способности, роли в патогенезе и морфогенезе туберкулезного процесса». В Пере-Жоан Кардона (ред.). Понимание туберкулеза - анализ происхождения патогенности Mycobacterium Tuberculosis. Хорватия: InTech. С. 179–206. ISBN 978-953-307-942-4.
внешняя ссылка
- Эпителиоид + клетки в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
| Этот клеточная биология статья - это заглушка. Вы можете помочь Википедии расширяя это. |